

Multi-modality in gene regulatory networks with slow promoter kinetics
- PMID: 30779734
- PMCID: PMC6396950
- DOI: 10.1371/journal.pcbi.1006784
Multi-modality in gene regulatory networks with slow promoter kinetics
Abstract
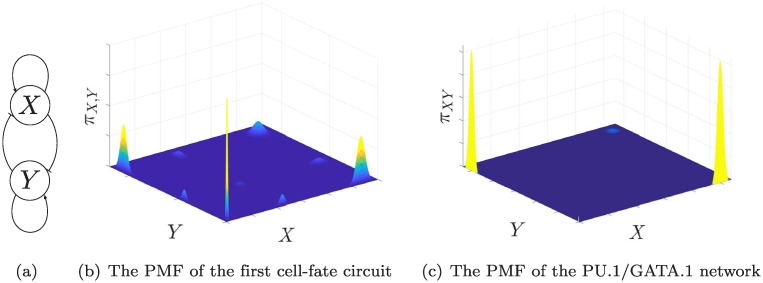
Phenotypical variability in the absence of genetic variation often reflects complex energetic landscapes associated with underlying gene regulatory networks (GRNs). In this view, different phenotypes are associated with alternative states of complex nonlinear systems: stable attractors in deterministic models or modes of stationary distributions in stochastic descriptions. We provide theoretical and practical characterizations of these landscapes, specifically focusing on stochastic Slow Promoter Kinetics (SPK), a time scale relevant when transcription factor binding and unbinding are affected by epigenetic processes like DNA methylation and chromatin remodeling. In this case, largely unexplored except for numerical simulations, adiabatic approximations of promoter kinetics are not appropriate. In contrast to the existing literature, we provide rigorous analytic characterizations of multiple modes. A general formal approach gives insight into the influence of parameters and the prediction of how changes in GRN wiring, for example through mutations or artificial interventions, impact the possible number, location, and likelihood of alternative states. We adapt tools from the mathematical field of singular perturbation theory to represent stationary distributions of Chemical Master Equations for GRNs as mixtures of Poisson distributions and obtain explicit formulas for the locations and probabilities of metastable states as a function of the parameters describing the system. As illustrations, the theory is used to tease out the role of cooperative binding in stochastic models in comparison to deterministic models, and applications are given to various model systems, such as toggle switches in isolation or in communicating populations, a synthetic oscillator, and a trans-differentiation network.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
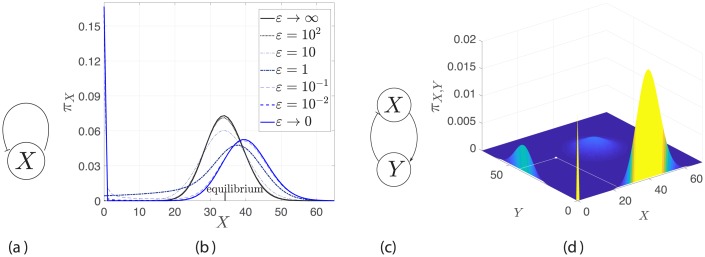
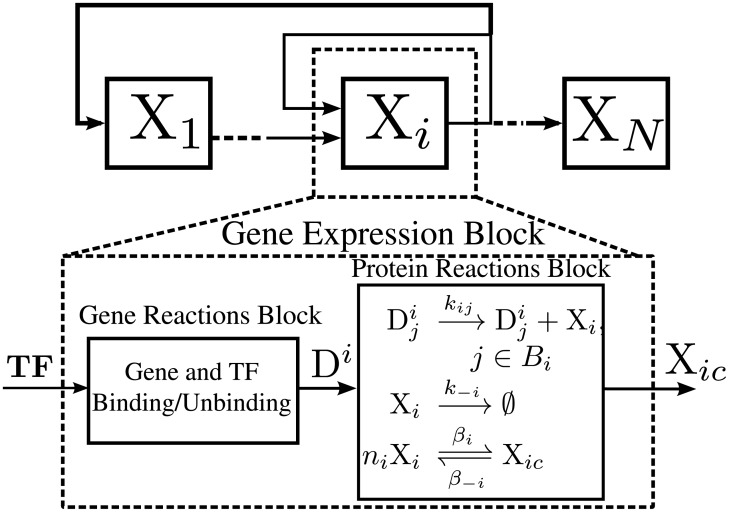
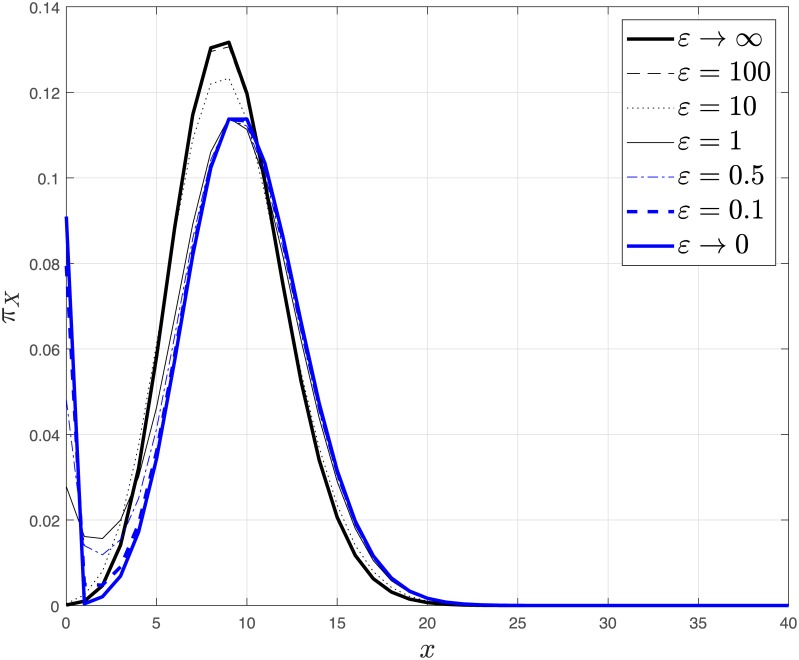
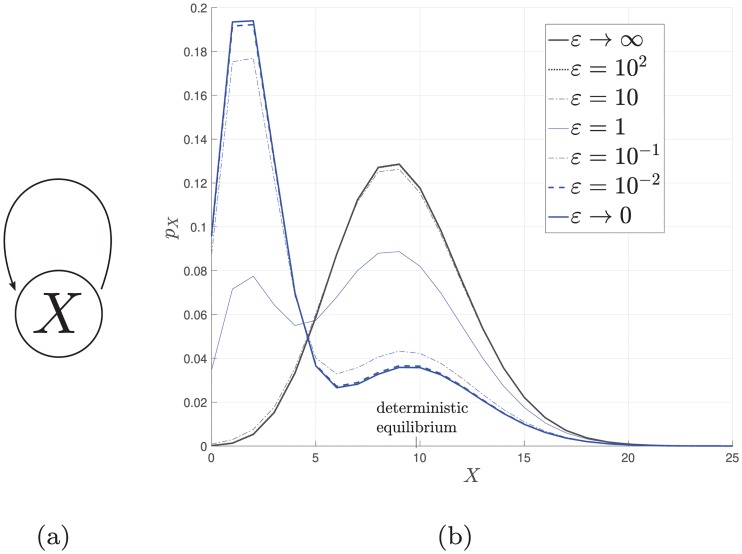
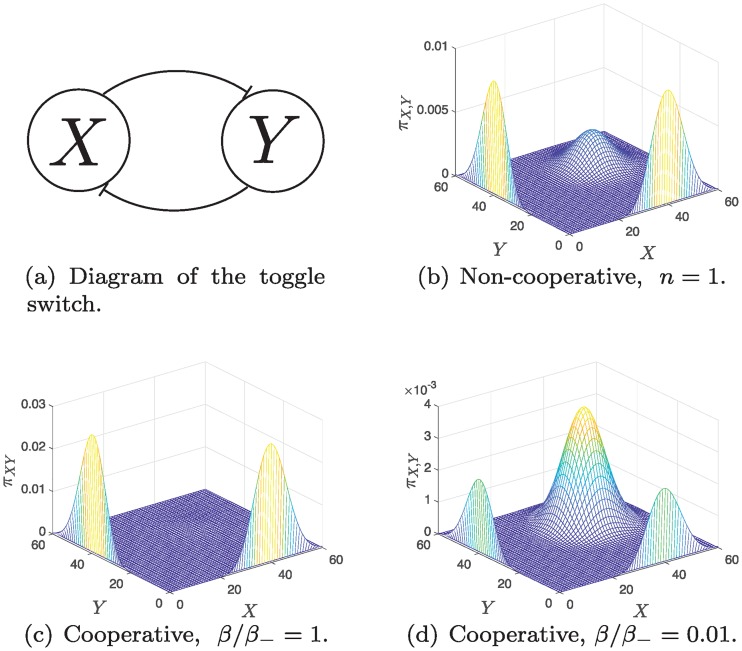
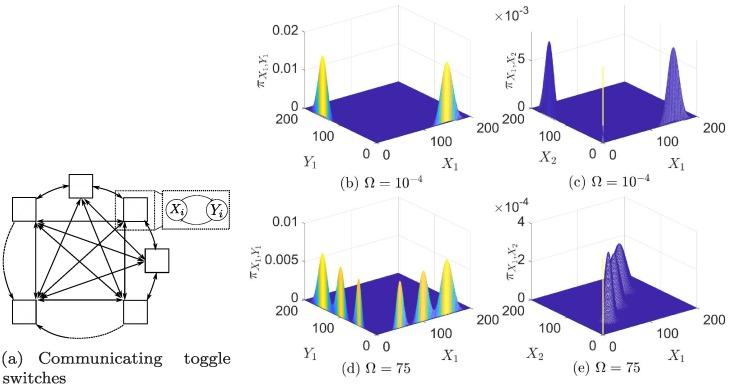
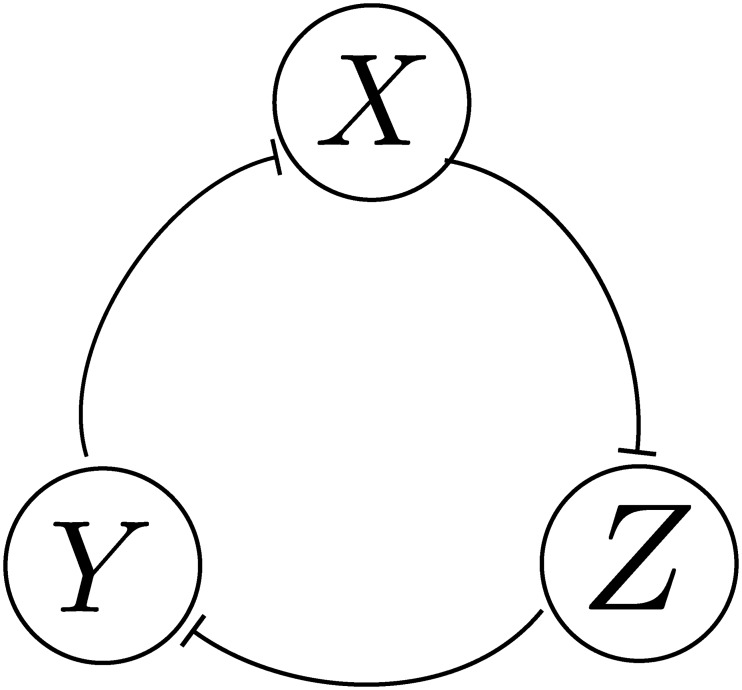
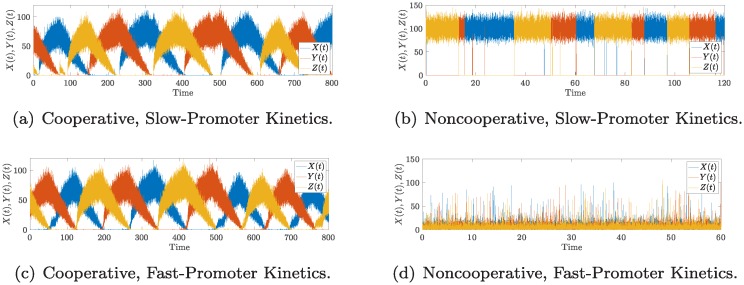
References
-
- Alon U. An Introduction to Systems Biology. CRC press; 2006.
-
- Del Vecchio D, Murray RM. Biomolecular Feedback Systems. Princeton Univ Press; 2015.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
