

Evolution of the multi-tRNA synthetase complex and its role in cancer
- PMID: 30782841
- PMCID: PMC6462501
- DOI: 10.1074/jbc.REV118.002958
Evolution of the multi-tRNA synthetase complex and its role in cancer
Abstract
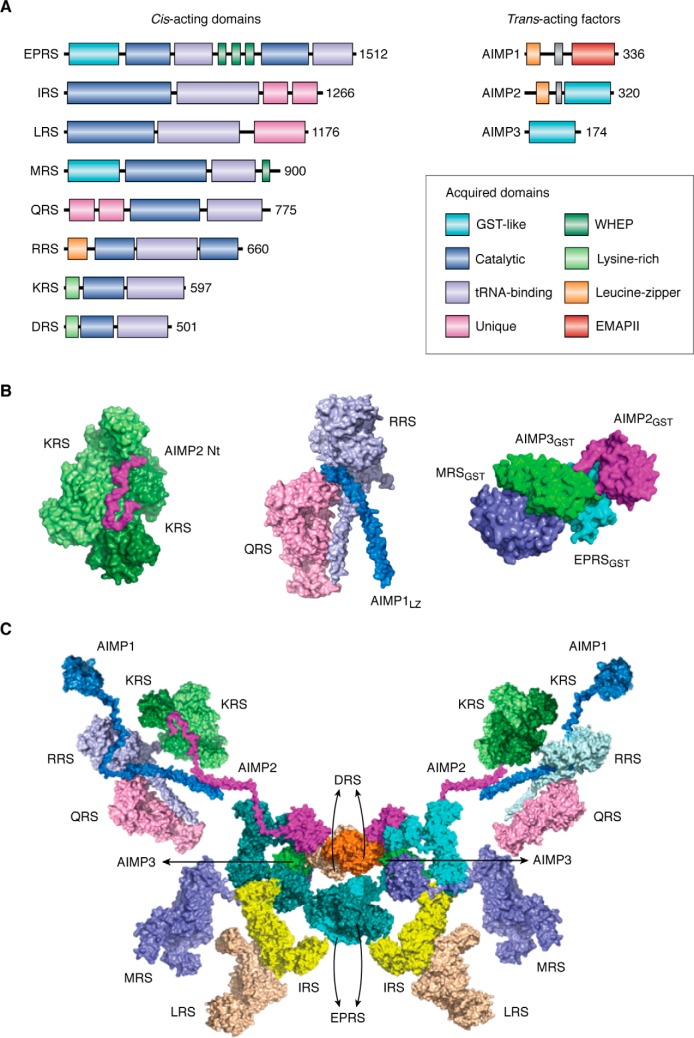
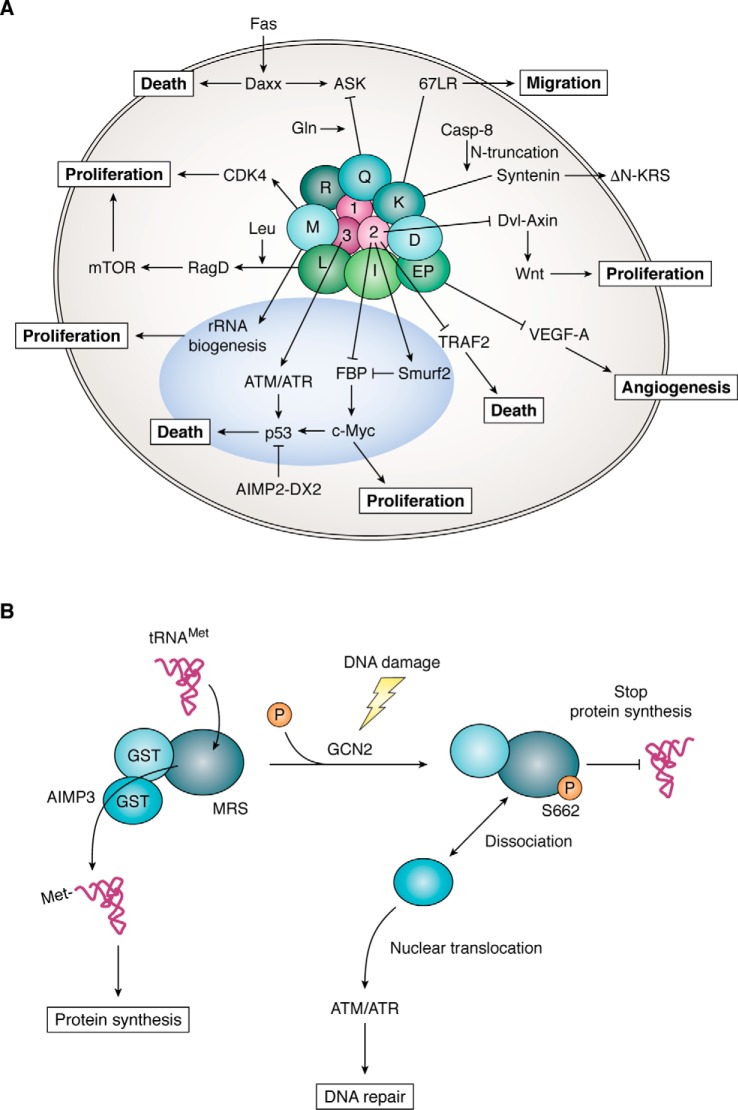
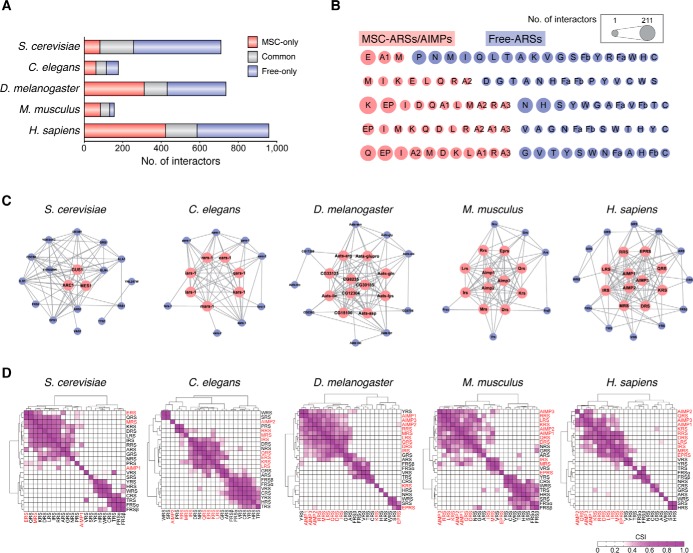
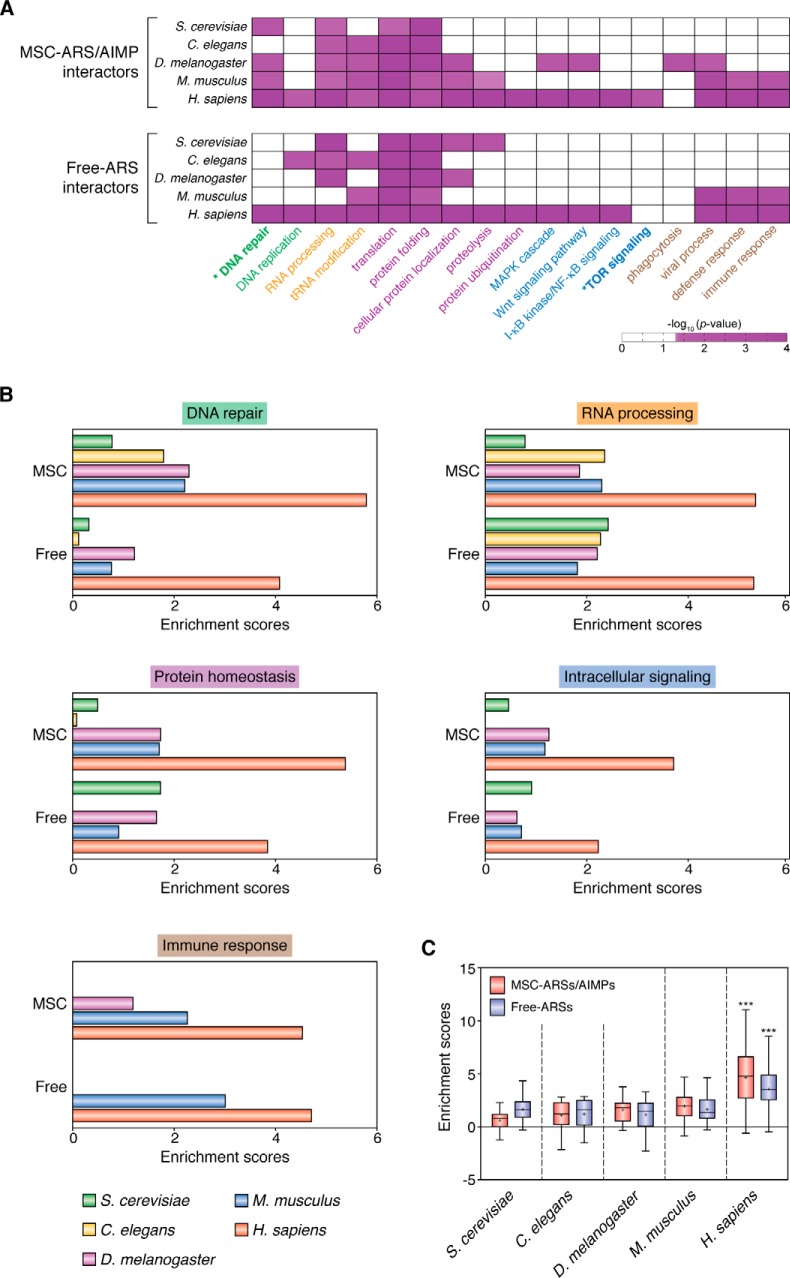
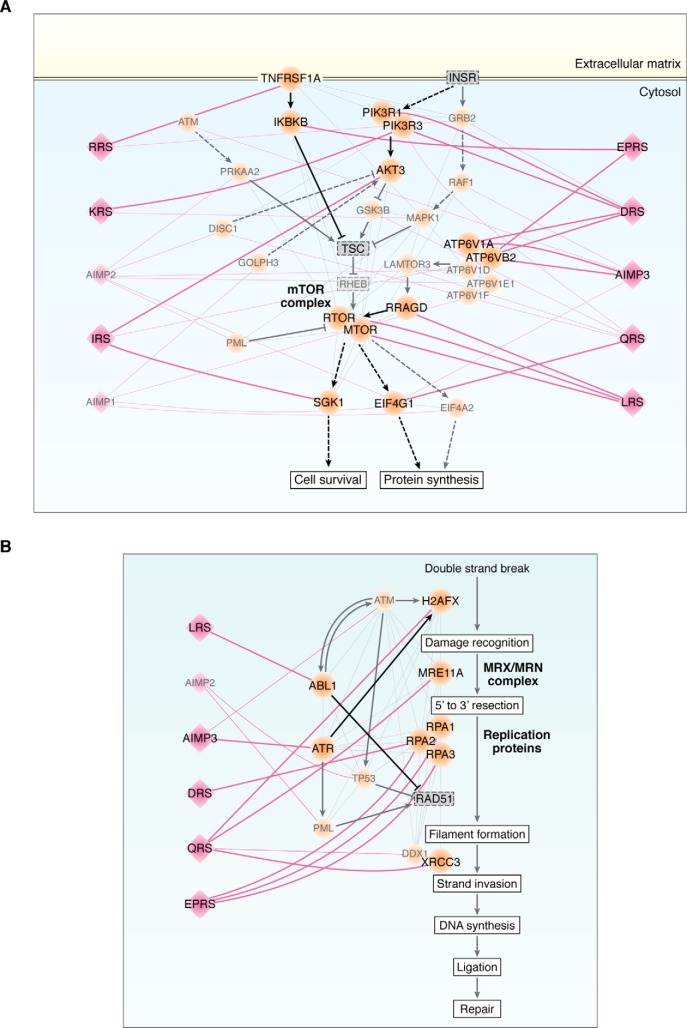
Aminoacyl-tRNA synthetases (ARSs) are enzymes that ligate their cognate amino acids to tRNAs for protein synthesis. However, recent studies have shown that their functions are expanded beyond protein synthesis through the interactions with diverse cellular factors. In this review, we discuss how ARSs have evolved to expand and control their functions by forming protein assemblies. We particularly focus on a macromolecular ARS complex in eukaryotes, named multi-tRNA synthetase complex (MSC), which is proposed to provide a channel through which tRNAs reach bound ARSs to receive their cognate amino acid and transit further to the translation machinery. Approximately half of the ARSs assemble into the MSC through cis-acting noncatalytic domains attached to their catalytic domains and trans-acting factors. Evolution of the MSC included its functional expansion, during which the MSC interaction network was augmented by additional cellular pathways present in higher eukaryotes. We also discuss MSC components that could be functionally involved in the pathophysiology of tumorigenesis. For example, the activities of some trans-acting factors have tumor-suppressing effects or maintain DNA integrity and are functionally compromised in cancer. On the basis of Gene Ontology analyses, we propose that the regulatory activities of the MSC-associated ARSs mainly converge on five biological processes, including mammalian target of rapamycin (mTOR) and DNA repair pathways. Future studies are needed to investigate how the MSC-associated and free-ARSs interact with each other and other factors in the control of multiple cellular pathways, and how aberrant or disrupted interactions in the MSC can cause disease.
Keywords: Aminoacyl-tRNA synthetases; Cancer; aminoacyl tRNA synthetase; cancer biology; intracellular processing; multi-tRNA synthetase complex; network analysis; pathology; protein synthesis; protein–protein interaction.
© 2019 Hyeon et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
-
- Swanson R., Hoben P., Sumner-Smith M., Uemura H., Watson L., and Söll D. (1988) Accuracy of in vivo aminoacylation requires proper balance of tRNA and aminoacyl-tRNA synthetase. Science 242, 1548–1551 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
