

Alternative Enzyme Protection Assay To Overcome the Drawbacks of the Gentamicin Protection Assay for Measuring Entry and Intracellular Survival of Staphylococci
- PMID: 30782857
- PMCID: PMC6479035
- DOI: 10.1128/IAI.00119-19
Alternative Enzyme Protection Assay To Overcome the Drawbacks of the Gentamicin Protection Assay for Measuring Entry and Intracellular Survival of Staphylococci
Abstract
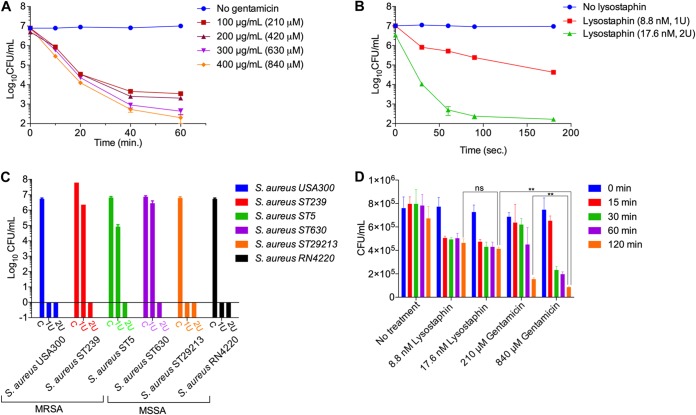
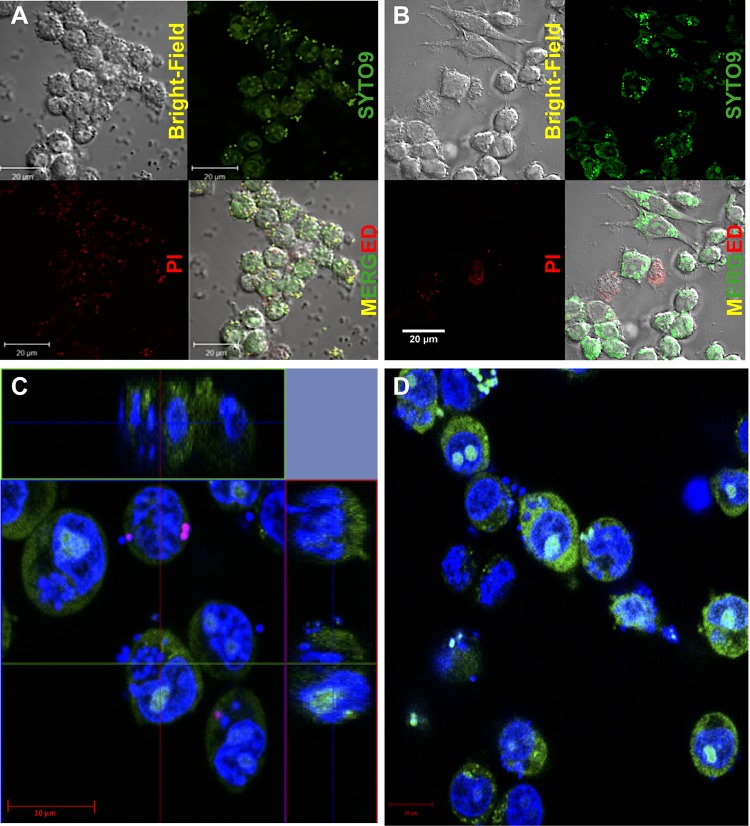
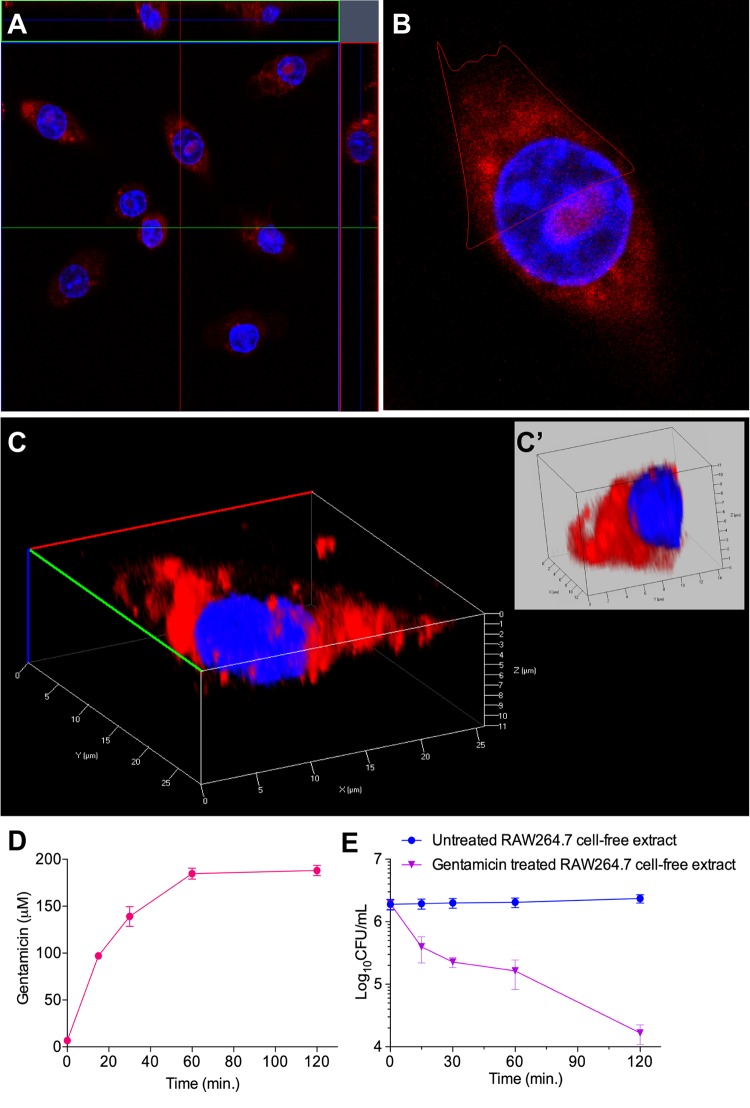
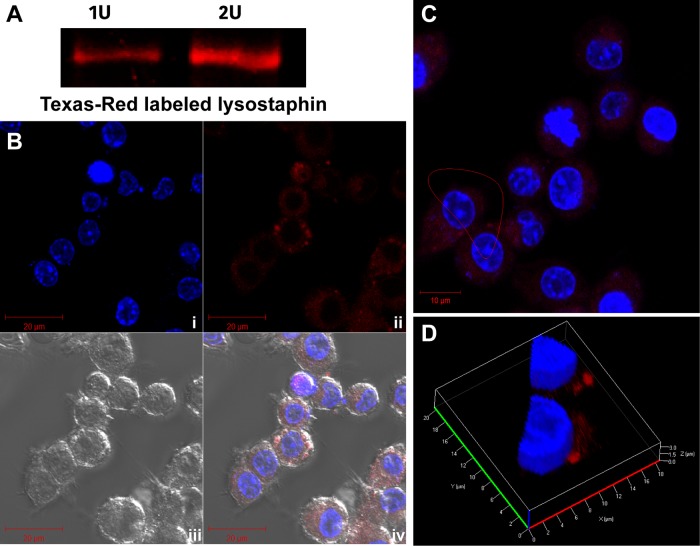
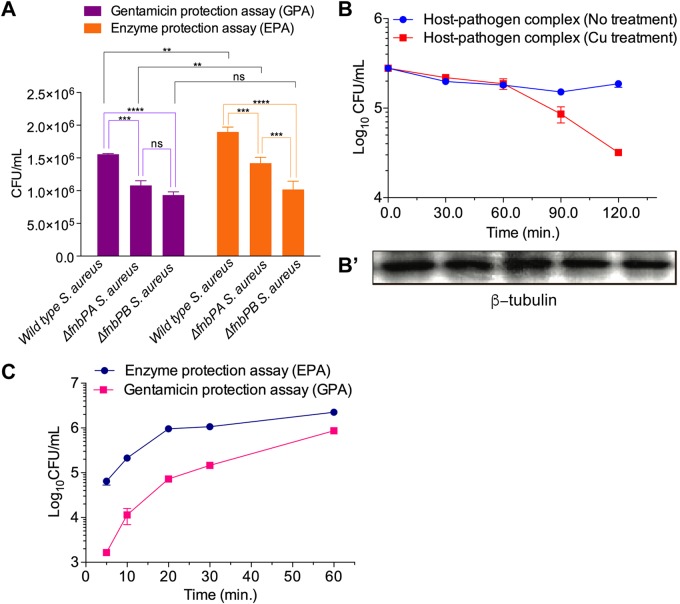
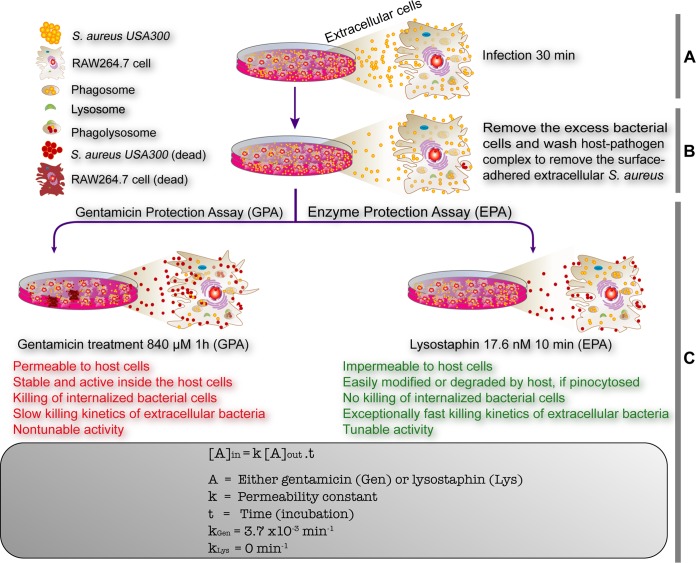
Precise enumeration of living intracellular bacteria is the key step to estimate the invasion potential of pathogens and host immune responses to understand the mechanism and kinetics of bacterial pathogenesis. Therefore, quantitative assessment of host-pathogen interactions is essential for development of novel antibacterial therapeutics for infectious disease. The gentamicin protection assay (GPA) is the most widely used method for these estimations by counting the CFU of intracellular living pathogens. Here, we assess the longstanding drawbacks of the GPA by employing an antistaphylococcal endopeptidase as a bactericidal agent to kill extracellular Staphylococcus aureus We found that the difference between the two methods for the recovery of intracellular CFU of S. aureus was about 5 times. We prove that the accurate number of intracellular CFU could not be precisely determined by the GPA due to the internalization of gentamicin into host cells during extracellular bacterial killing. We further demonstrate that lysostaphin-mediated extracellular bacterial clearance has advantages for measuring the kinetics of bacterial internalization on a minute time scale due to the fast and tunable activity and the inability of protein to permeate the host cell membrane. From these results, we propose that accurate quantification of intracellular bacteria and measurement of internalization kinetics can be achieved by employing enzyme-mediated killing of extracellular bacteria (enzyme protection assay [EPA]) rather than the host-permeative drug gentamicin, which is known to alter host physiology.
Keywords: Staphylococcus aureus; bacteria; enzyme protection assay; gentamicin protection assay; host; lysostaphin.
Copyright © 2019 American Society for Microbiology.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
