

An FBXW7-ZEB2 axis links EMT and tumour microenvironment to promote colorectal cancer stem cells and chemoresistance
- PMID: 30783098
- PMCID: PMC6381143
- DOI: 10.1038/s41389-019-0125-3
An FBXW7-ZEB2 axis links EMT and tumour microenvironment to promote colorectal cancer stem cells and chemoresistance
Abstract
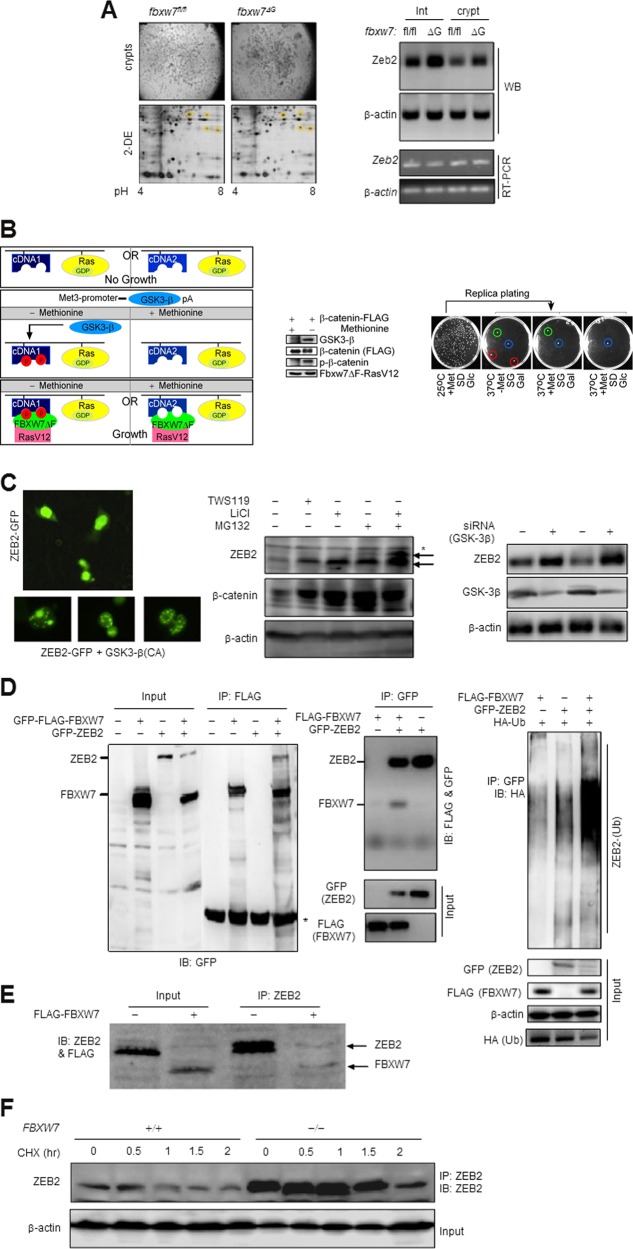
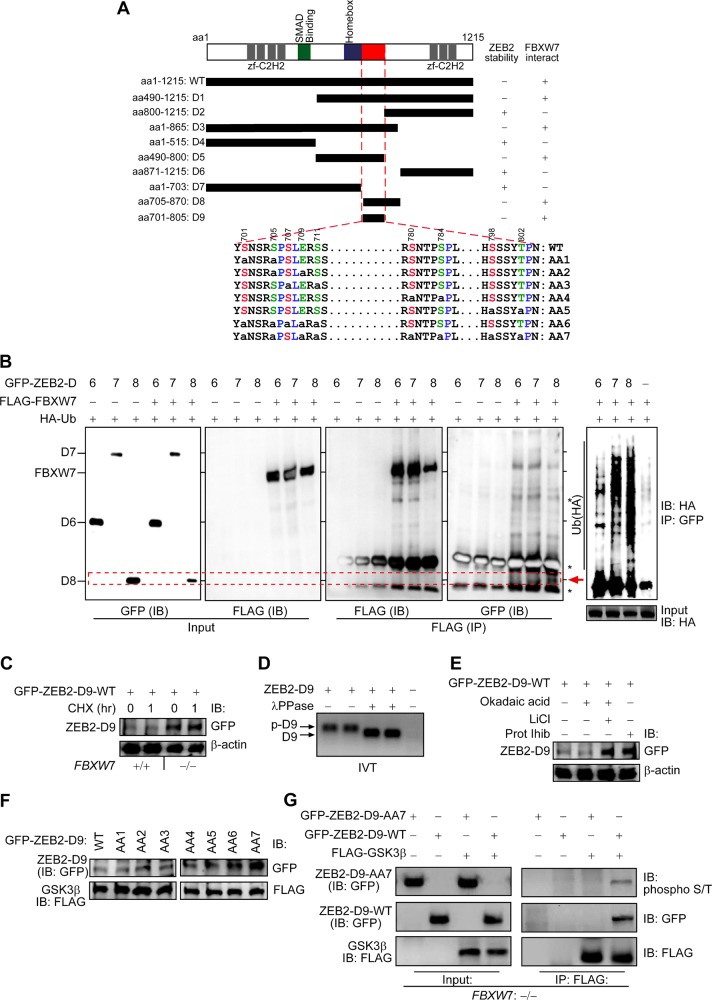
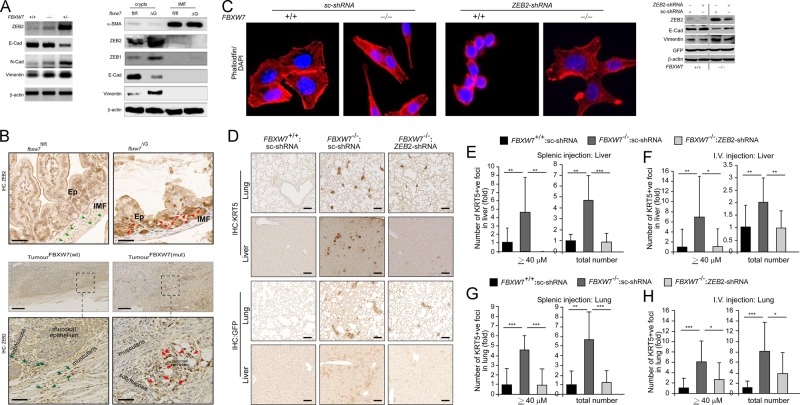
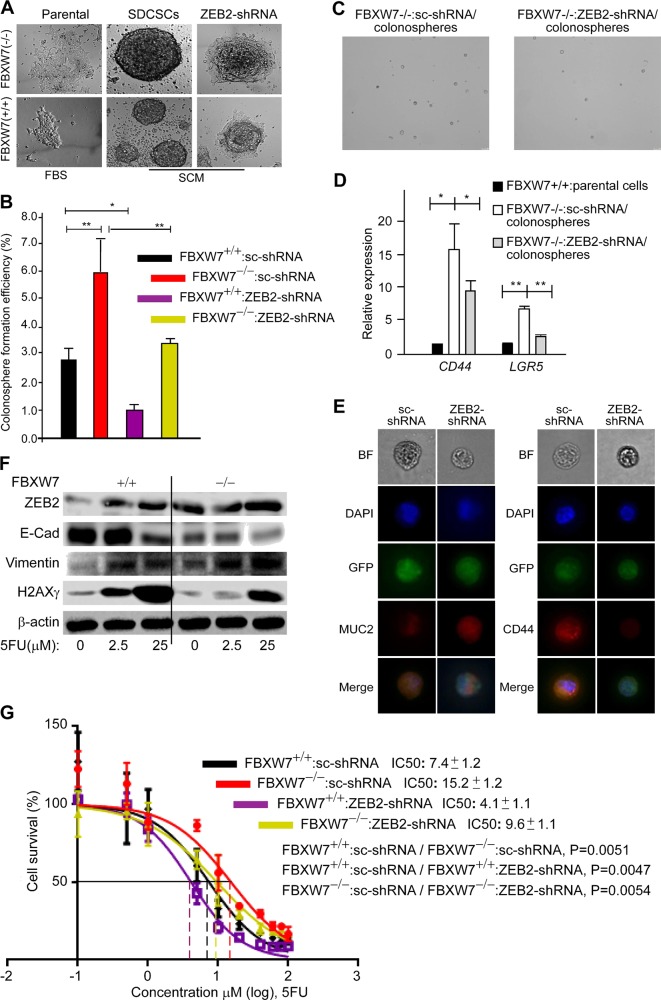
Colorectal cancer (CRC) patients develop recurrence after chemotherapy owing to the survival of stem cell-like cells referred to as cancer stem-like cells (CSCs). The origin of CSCs is linked to the epithelial-mesenchymal transition (EMT) process. Currently, it remains poorly understood how EMT programmes enable CSCs residing in the tumour microenvironment to escape the effects of chemotherapy. This study identifies a key molecular pathway that is responsible for the formation of drug-resistant CSC populations. Using a modified yeast-2-hybrid system and 2D gel-based proteomics methods, we show that the E3-ubiquitin ligase FBXW7 directly binds and degrades the EMT-inducing transcription factor ZEB2 in a phosphorylation-dependent manner. Loss of FBXW7 induces an EMT that can be effectively reversed by knockdown of ZEB2. The FBXW7-ZEB2 axis regulates such important cancer cell features, as stemness/dedifferentiation, chemoresistance and cell migration in vitro, ex vivo and in animal models of metastasis. High expression of ZEB2 in cancer tissues defines the reduced ZEB2 expression in the cancer-associated stroma in patients and in murine intestinal organoids, demonstrating a tumour-stromal crosstalk that modulates a niche and EMT activation. Our study thus uncovers a new molecular mechanism, by which the CRC cells display differences in resistance to chemotherapy and metastatic potential.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
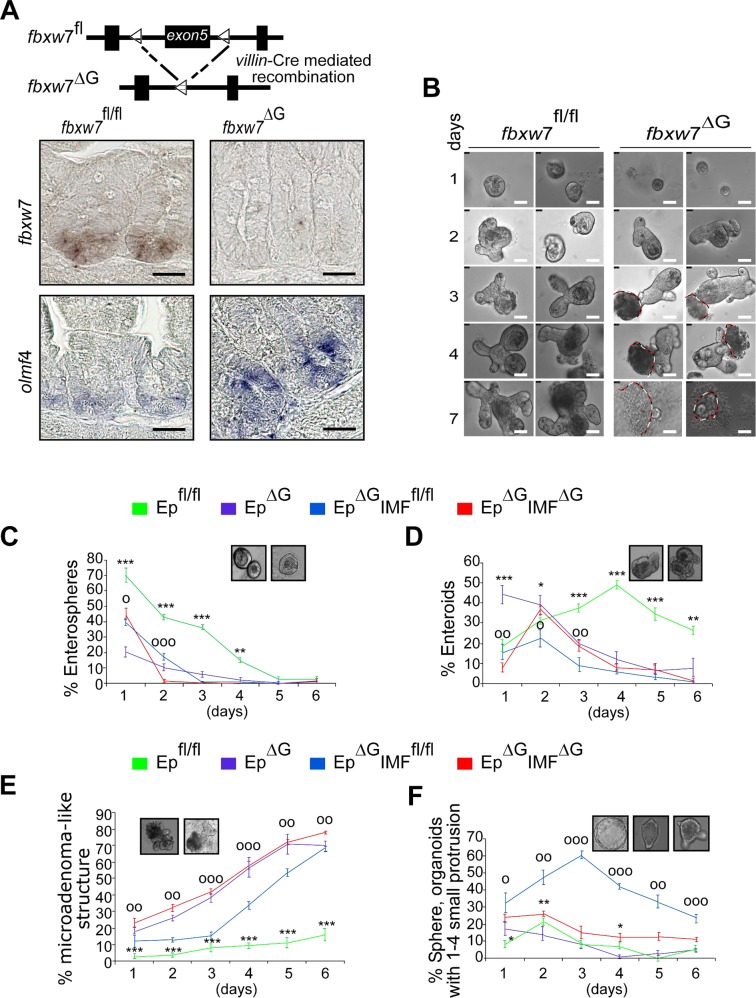
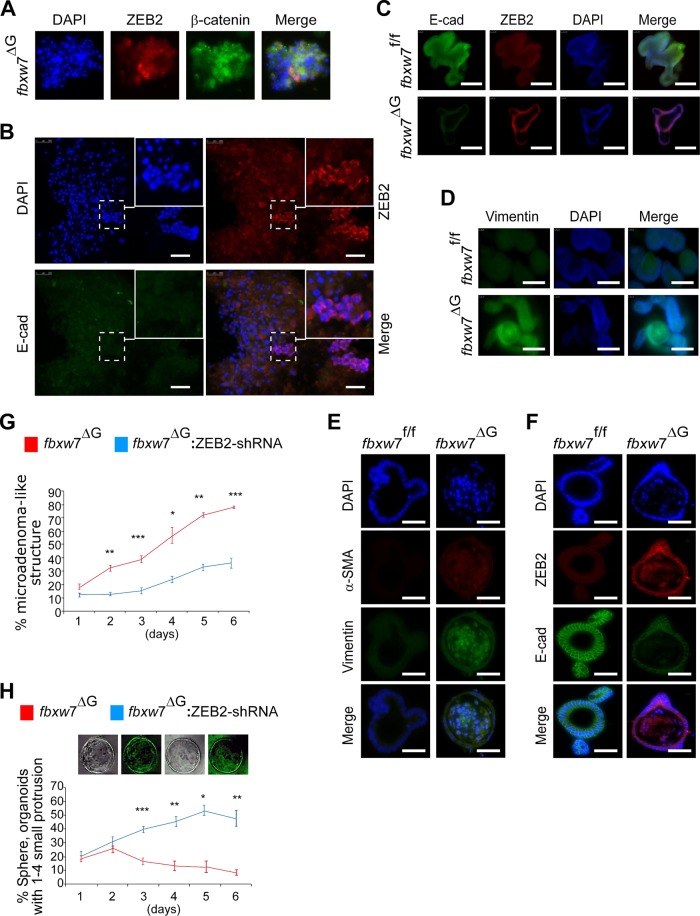
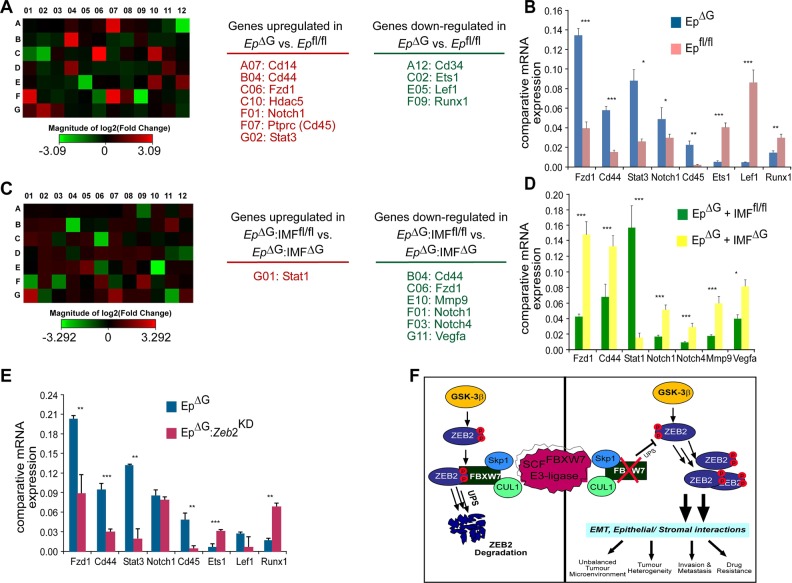
References
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
