

Protein S-nitrosylation in programmed cell death in plants
- PMID: 30783684
- PMCID: PMC11105606
- DOI: 10.1007/s00018-019-03045-0
Protein S-nitrosylation in programmed cell death in plants
Abstract
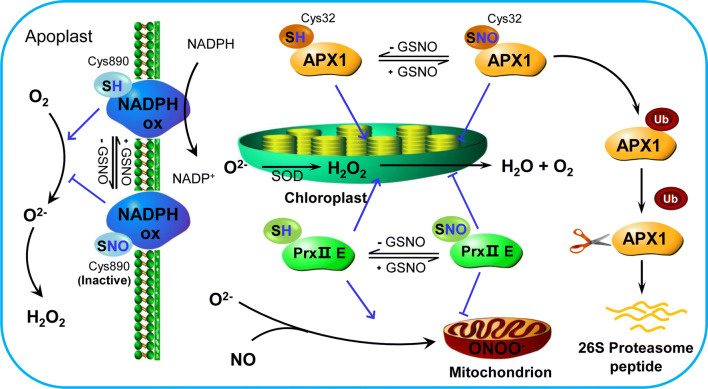
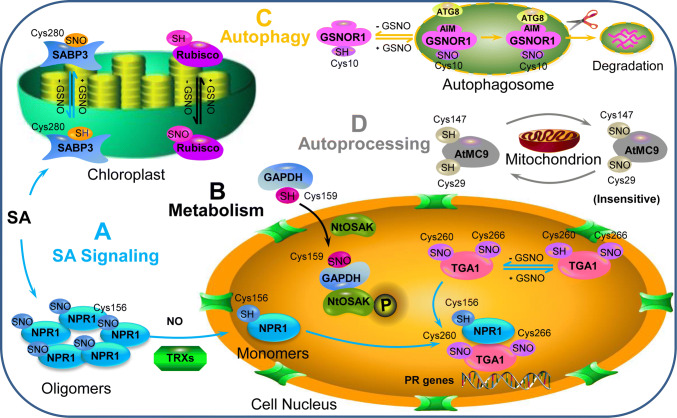
Programmed cell death (PCD) is associated with different phases of plant life and provides resistance to different kinds of biotic or abiotic stress. The redox molecule nitric oxide (NO) is usually produced during the stress response and exerts dual effects on PCD regulation. S-nitrosylation, which NO attaches to the cysteine thiol of proteins, is a vital posttranslational modification and is considered as an essential way for NO to regulate cellular redox signaling. In recent years, a great number of proteins have been identified as targets of S-nitrosylation in plants, especially during PCD. S-nitrosylation can directly affect plant PCD positively or negatively, mainly by regulating the activity of cell death-related enzymes or reconstructing the conformation of several functional proteins. Here, we summarized S-nitrosylated proteins that are involved in PCD and provide insight into how S-nitrosylation can regulate plant PCD. In addition, both the importance and challenges of future works on S-nitrosylation in plant PCD are highlighted.
Keywords: Plants; Programmed cell death; Protein S-nitrosylation.
Figures
Similar articles
-
Protein S-nitrosylation under abiotic stress: Role and mechanism.Plant Physiol Biochem. 2024 Feb;207:108329. doi: 10.1016/j.plaphy.2023.108329. Epub 2024 Jan 4. Plant Physiol Biochem. 2024. PMID: 38184883 Review.
-
Protein S-nitrosylation in plants: photorespiratory metabolism and NO signaling.Sci Signal. 2011 Jan 4;4(154):jc1. doi: 10.1126/scisignal.2001404. Sci Signal. 2011. PMID: 21205936 Review.
-
Protein S-Nitrosylation in plants: Current progresses and challenges.J Integr Plant Biol. 2019 Dec;61(12):1206-1223. doi: 10.1111/jipb.12780. Epub 2019 Mar 27. J Integr Plant Biol. 2019. PMID: 30663237 Review.
-
The roles of protein S-nitrosylation in regulating the growth and development of plants: A review.Int J Biol Macromol. 2025 May;307(Pt 3):142204. doi: 10.1016/j.ijbiomac.2025.142204. Epub 2025 Mar 17. Int J Biol Macromol. 2025. PMID: 40107544 Review.
-
Sub-proteome S-nitrosylation analysis in Brassica juncea hints at the regulation of Brassicaceae specific as well as other vital metabolic pathway(s) by nitric oxide and suggests post-translational modifications cross-talk.Nitric Oxide. 2014 Dec 1;43:97-111. doi: 10.1016/j.niox.2014.08.010. Epub 2014 Aug 28. Nitric Oxide. 2014. PMID: 25175897 Review.
Cited by
-
qPTMplants: an integrative database of quantitative post-translational modifications in plants.Nucleic Acids Res. 2022 Jan 7;50(D1):D1491-D1499. doi: 10.1093/nar/gkab945. Nucleic Acids Res. 2022. PMID: 34718741 Free PMC article.
-
Involvement of Nitric Oxide and Melatonin Enhances Cadmium Resistance of Tomato Seedlings through Regulation of the Ascorbate-Glutathione Cycle and ROS Metabolism.Int J Mol Sci. 2023 May 31;24(11):9526. doi: 10.3390/ijms24119526. Int J Mol Sci. 2023. PMID: 37298477 Free PMC article.
-
Role of protein S-nitrosylation in plant growth and development.Plant Cell Rep. 2024 Jul 30;43(8):204. doi: 10.1007/s00299-024-03290-z. Plant Cell Rep. 2024. PMID: 39080060 Review.
-
Dual Roles of GSNOR1 in Cell Death and Immunity in Tetraploid Nicotiana tabacum.Front Plant Sci. 2021 Feb 10;12:596234. doi: 10.3389/fpls.2021.596234. eCollection 2021. Front Plant Sci. 2021. PMID: 33643341 Free PMC article.
-
Regulation of Ascorbate-Glutathione Pathway in Mitigating Oxidative Damage in Plants under Abiotic Stress.Antioxidants (Basel). 2019 Sep 9;8(9):384. doi: 10.3390/antiox8090384. Antioxidants (Basel). 2019. PMID: 31505852 Free PMC article. Review.
References
-
- Locato V, Paradiso A, Sabetta W, De Gara L, de Pinto MC. Advances in botanical research. New York: Academic; 2016. Nitric Oxide-biol Ch and reactive oxygen species in PCD signaling; pp. 165–192.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
