

Filamentous Aggregates Are Fragmented by the Proteasome Holoenzyme
- PMID: 30784595
- PMCID: PMC6381791
- DOI: 10.1016/j.celrep.2019.01.096
Filamentous Aggregates Are Fragmented by the Proteasome Holoenzyme
Abstract
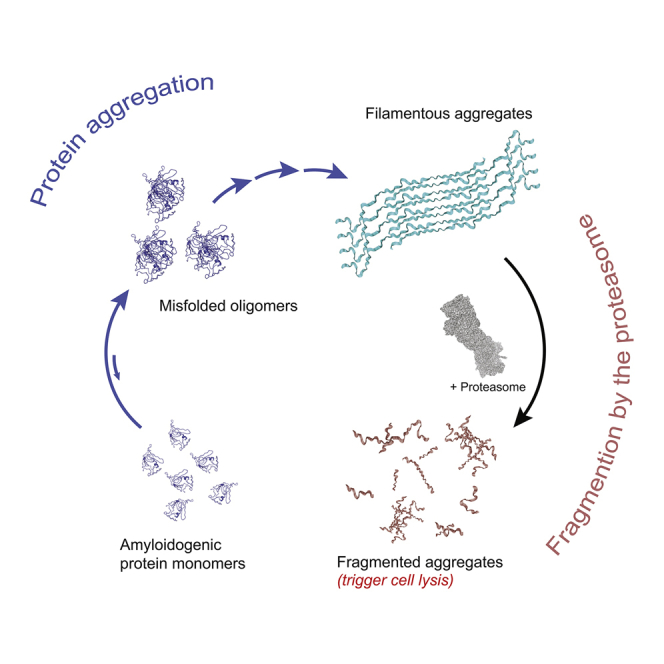
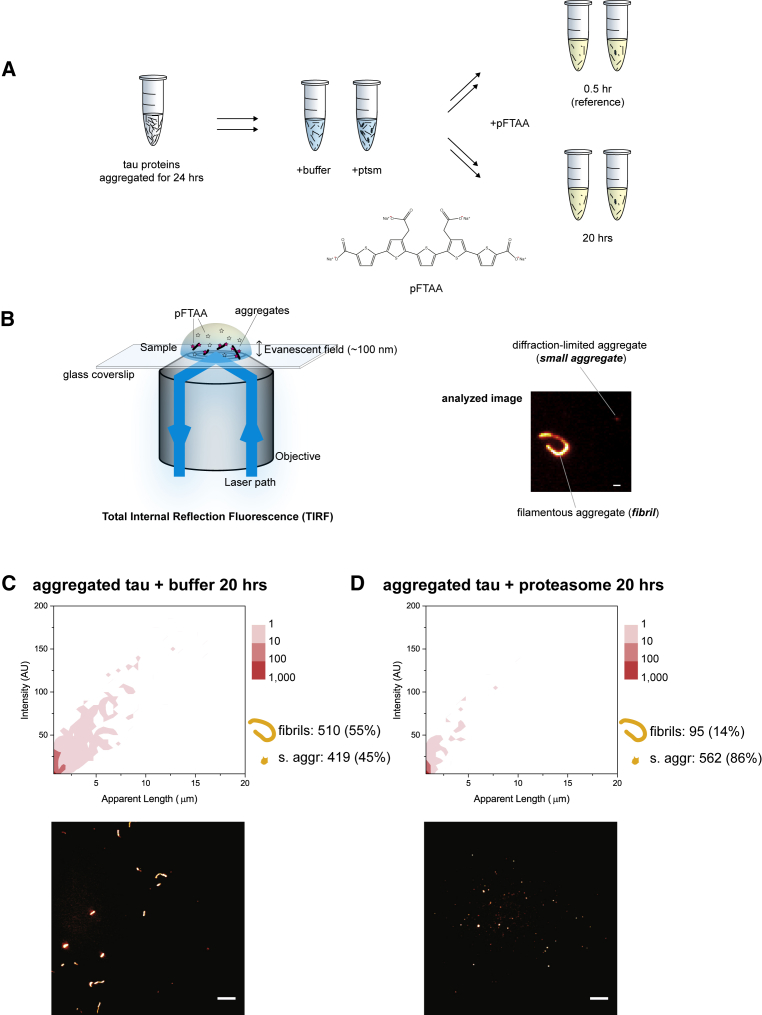
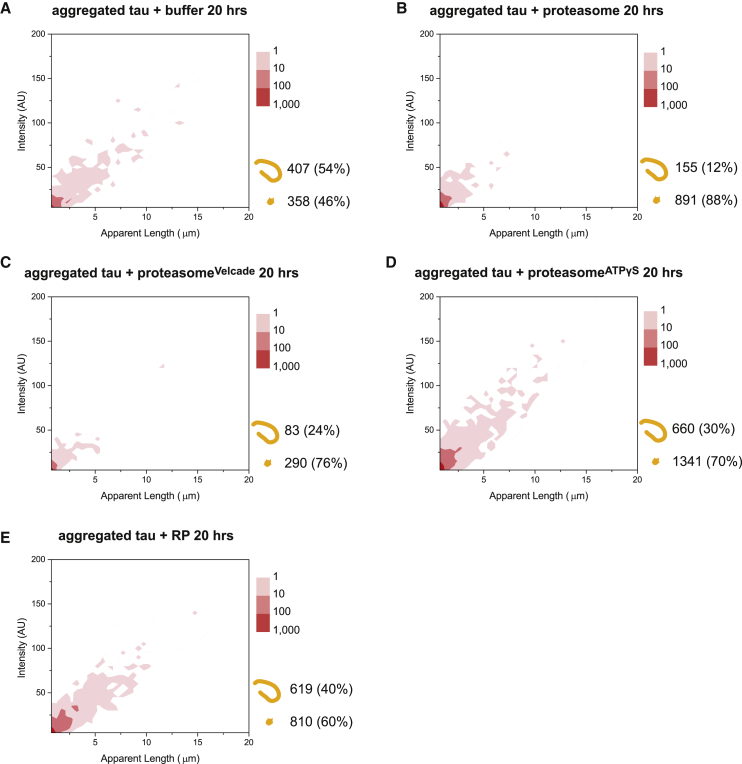
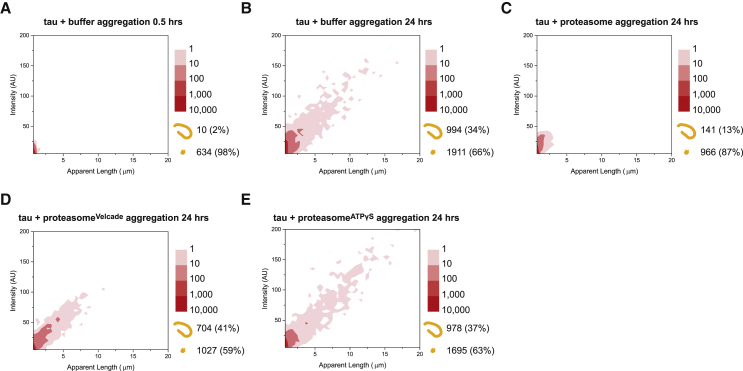
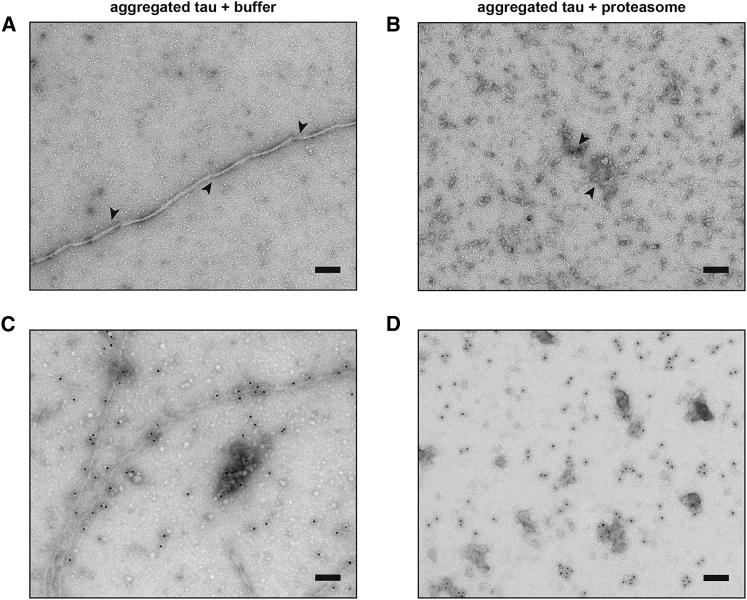
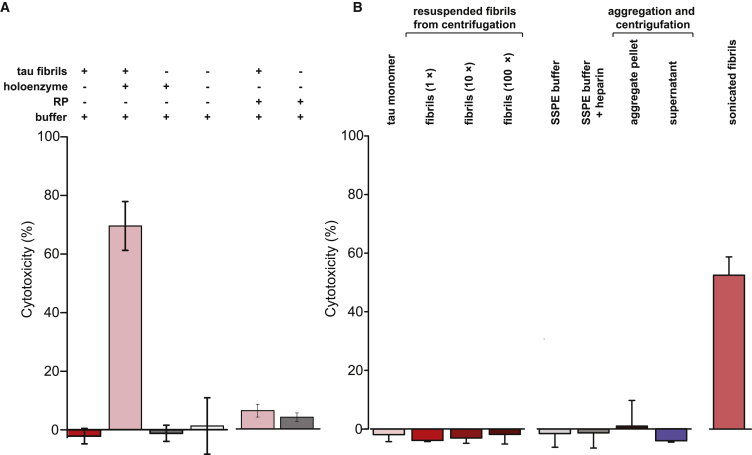
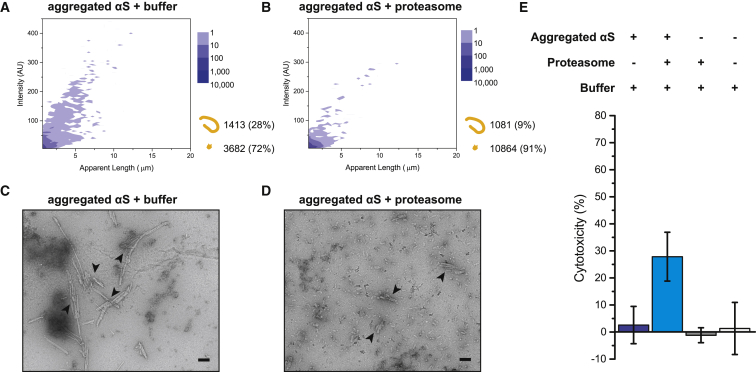
Filamentous aggregates (fibrils) are regarded as the final stage in the assembly of amyloidogenic proteins and are formed in many neurodegenerative diseases. Accumulation of aggregates occurs as a result of an imbalance between their formation and removal. Here we use single-aggregate imaging to show that large fibrils assembled from full-length tau are substrates of the 26S proteasome holoenzyme, which fragments them into small aggregates. Interestingly, although degradation of monomeric tau is not inhibited by adenosine 5'-(3-thiotriphosphate) (ATPγS), fibril fragmentation is predominantly dependent on the ATPase activity of the proteasome. The proteasome holoenzyme also targets fibrils assembled from α-synuclein, suggesting that its fibril-fragmenting function may be a general mechanism. The fragmented species produced by the proteasome shows significant toxicity to human cell lines compared with intact fibrils. Together, our results indicate that the proteasome holoenzyme possesses a fragmentation function that disassembles large fibrils into smaller and more cytotoxic species.
Keywords: alpha-synuclein; disaggregation; proteasome; protein aggregation; tau; total-internal reflection fluorescence microscopy.
Copyright © 2019 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Barrett P.J., Timothy Greenamyre J. Post-translational modification of α-synuclein in Parkinson’s disease. Brain Res. 2015;1628(Pt B):247–253. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
