

Acoustic Pattern Recognition and Courtship Songs: Insights from Insects
- PMID: 30786225
- PMCID: PMC6885008
- DOI: 10.1146/annurev-neuro-080317-061839
Acoustic Pattern Recognition and Courtship Songs: Insights from Insects
Abstract
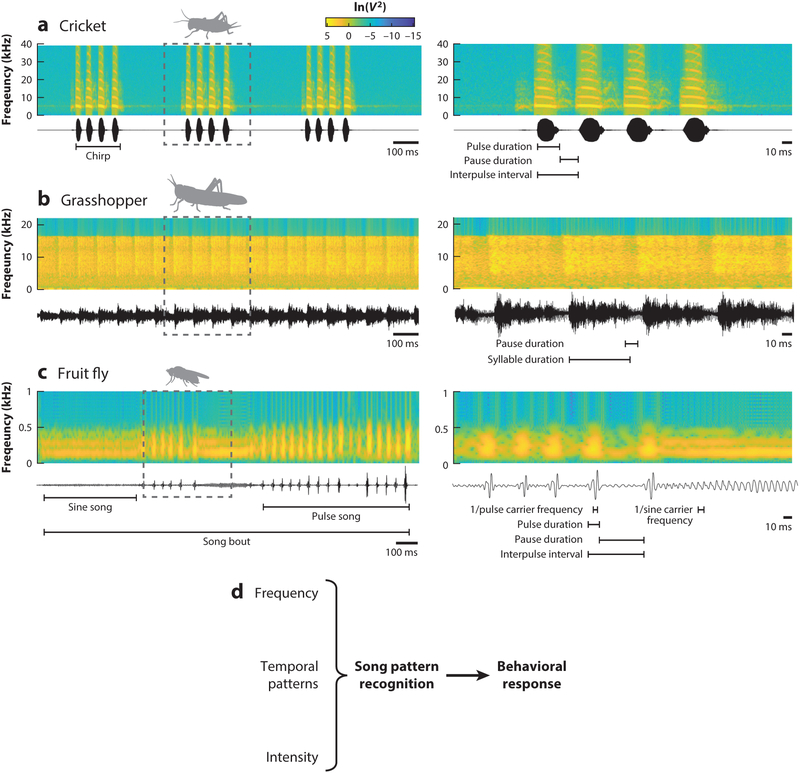
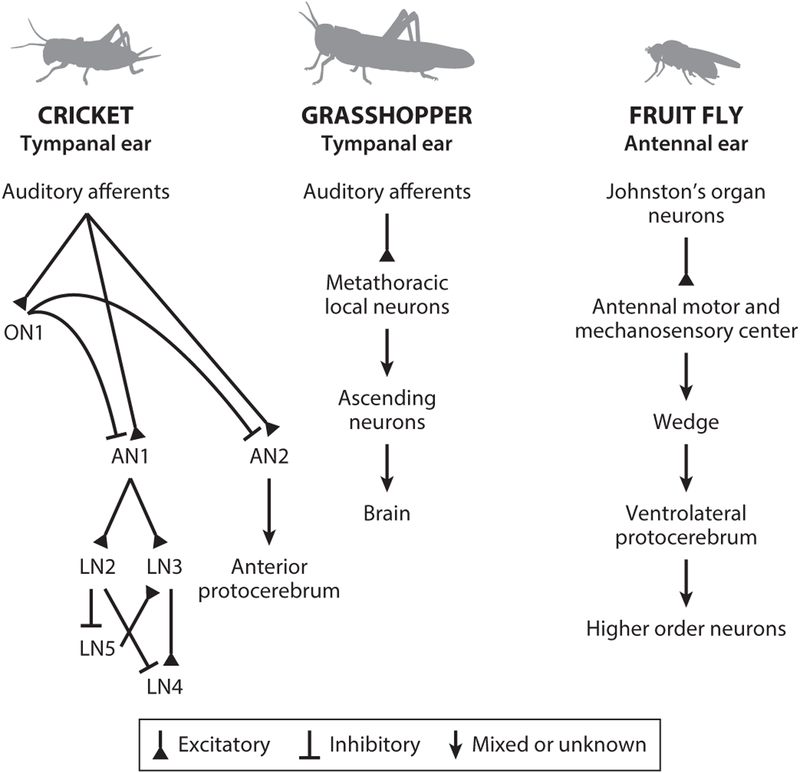
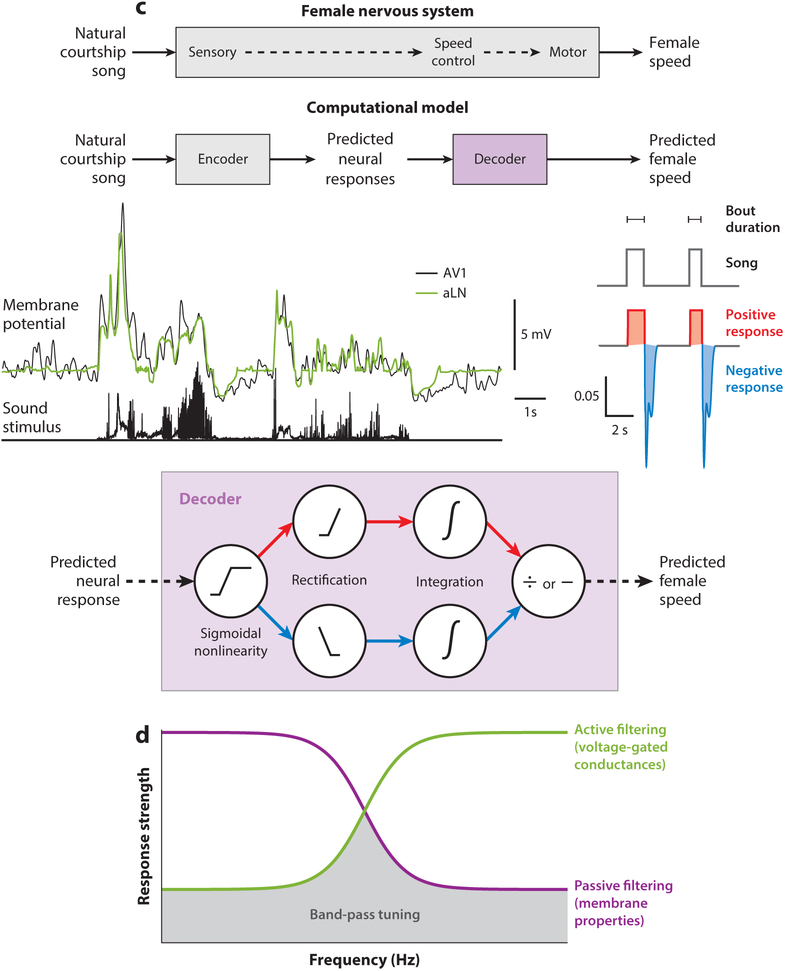
Across the animal kingdom, social interactions rely on sound production and perception. From simple cricket chirps to more elaborate bird songs, animals go to great lengths to communicate information critical for reproduction and survival via acoustic signals. Insects produce a wide array of songs to attract a mate, and the intended receivers must differentiate these calls from competing sounds, analyze the quality of the sender from spectrotemporal signal properties, and then determine how to react. Insects use numerically simple nervous systems to analyze and respond to courtship songs, making them ideal model systems for uncovering the neural mechanisms underlying acoustic pattern recognition. We highlight here how the combination of behavioral studies and neural recordings in three groups of insects-crickets, grasshoppers, and fruit flies-reveals common strategies for extracting ethologically relevant information from acoustic patterns and how these findings might translate to other systems.
Keywords: animal communication; auditory circuits; auditory processing; courtship songs; temporal pattern recognition.
Figures

References
-
- Alder TB, Rose GJ. 1998. Long-term temporal integration in the anuran auditory system. Nat. Neurosci 1(6):519–23 - PubMed
-
- Atkins G, Ligman S, Burghardt F, Stout JF. 1984. Changes in phonotaxis by the female cricket Acheta domesticusL. after killing identified acoustic interneurons. J. Comp. Physiol. A 154(6):795–804
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
