

The impact of proliferation-migration tradeoffs on phenotypic evolution in cancer
- PMID: 30787363
- PMCID: PMC6382810
- DOI: 10.1038/s41598-019-39636-x
The impact of proliferation-migration tradeoffs on phenotypic evolution in cancer
Abstract
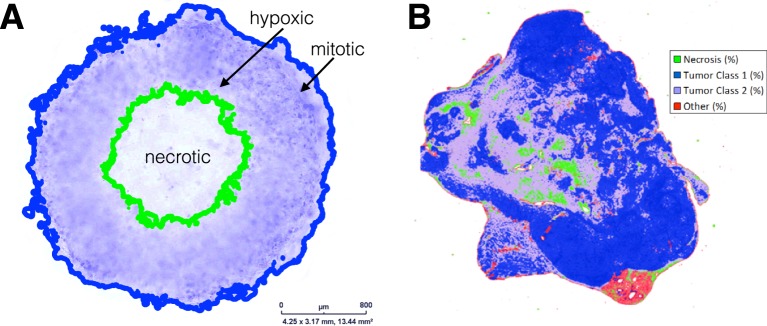
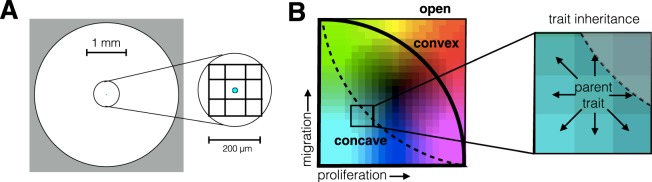
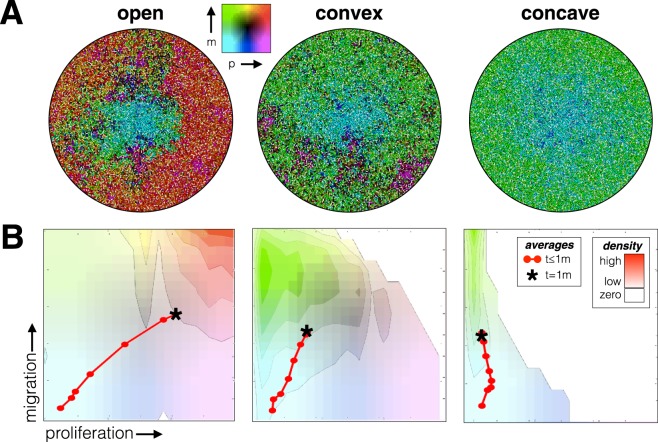
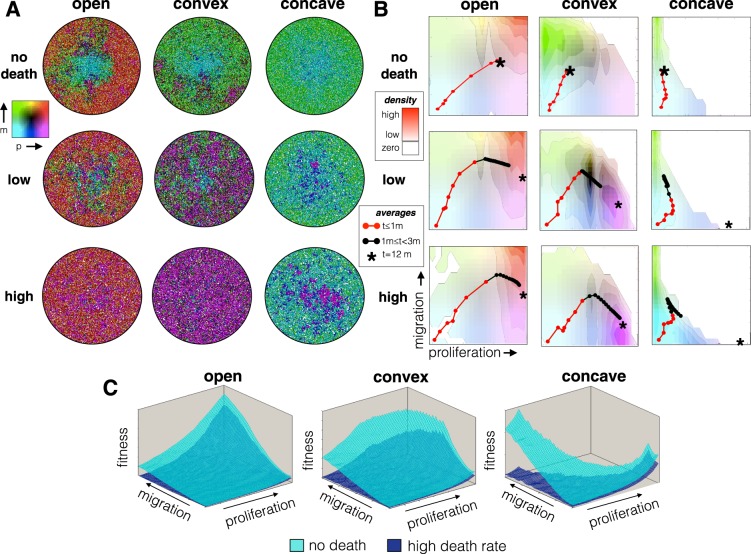
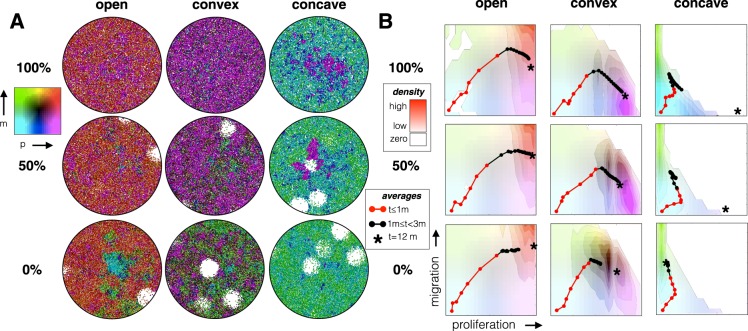
Tumors are not static masses of cells but dynamic ecosystems where cancer cells experience constant turnover and evolve fitness-enhancing phenotypes. Selection for different phenotypes may vary with (1) the tumor niche (edge or core), (2) cell turnover rates, (3) the nature of the tradeoff between traits, and (4) whether deaths occur in response to demographic or environmental stochasticity. Using a spatially-explicit agent-based model, we observe how two traits (proliferation rate and migration speed) evolve under different tradeoff conditions with different turnover rates. Migration rate is favored over proliferation at the tumor's edge and vice-versa for the interior. Increasing cell turnover rates slightly slows tumor growth but accelerates the rate of evolution for both proliferation and migration. The absence of a tradeoff favors ever higher values for proliferation and migration, while a convex tradeoff tends to favor proliferation, often promoting the coexistence of a generalist and specialist phenotype. A concave tradeoff favors migration at low death rates, but switches to proliferation at higher death rates. Mortality via demographic stochasticity favors proliferation, and environmental stochasticity favors migration. While all of these diverse factors contribute to the ecology, heterogeneity, and evolution of a tumor, their effects may be predictable and empirically accessible.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
