

BFSP1 C-terminal domains released by post-translational processing events can alter significantly the calcium regulation of AQP0 water permeability
- PMID: 30790544
- PMCID: PMC6713518
- DOI: 10.1016/j.exer.2019.02.001
BFSP1 C-terminal domains released by post-translational processing events can alter significantly the calcium regulation of AQP0 water permeability
Abstract
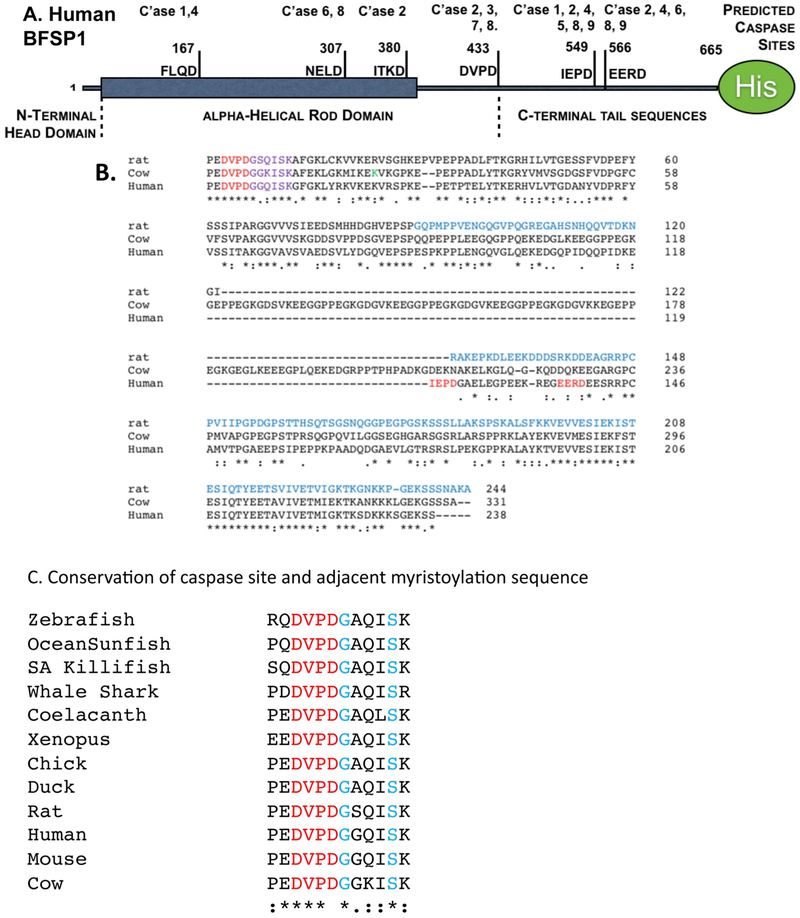
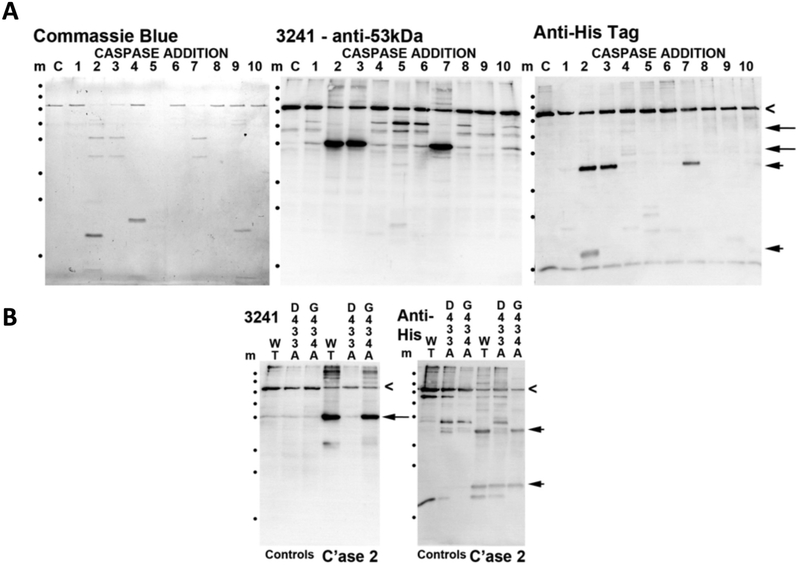
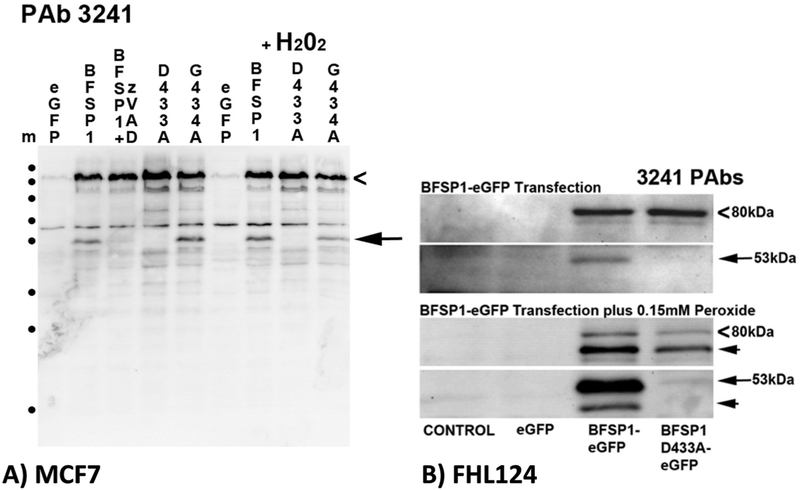
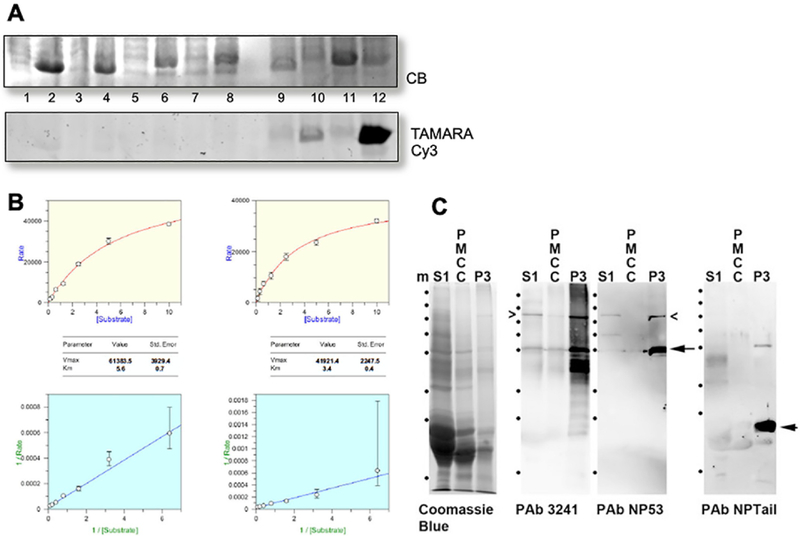
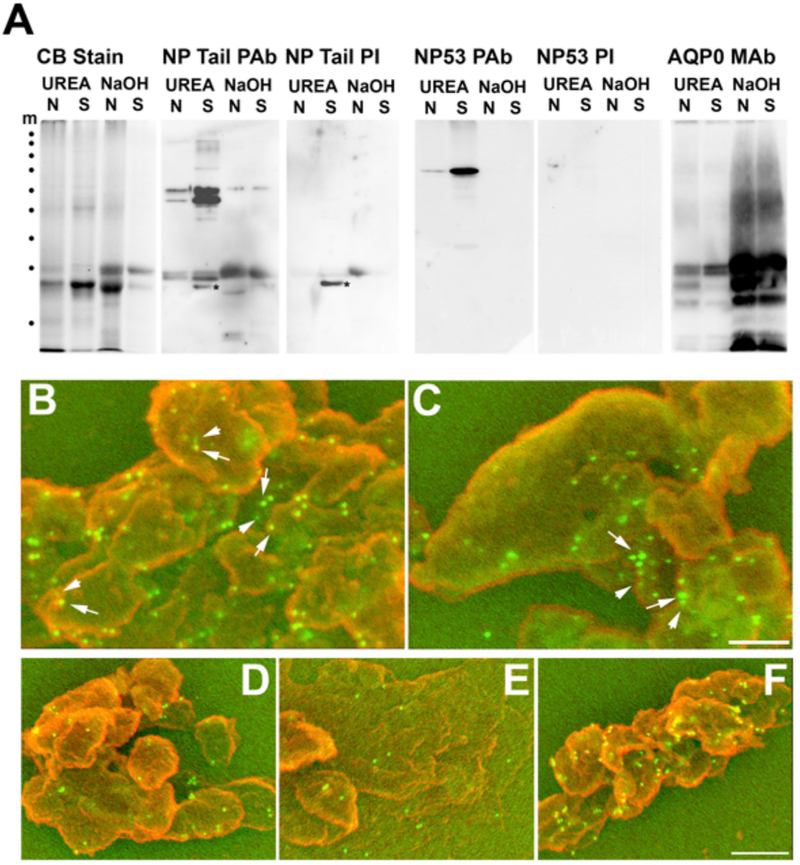
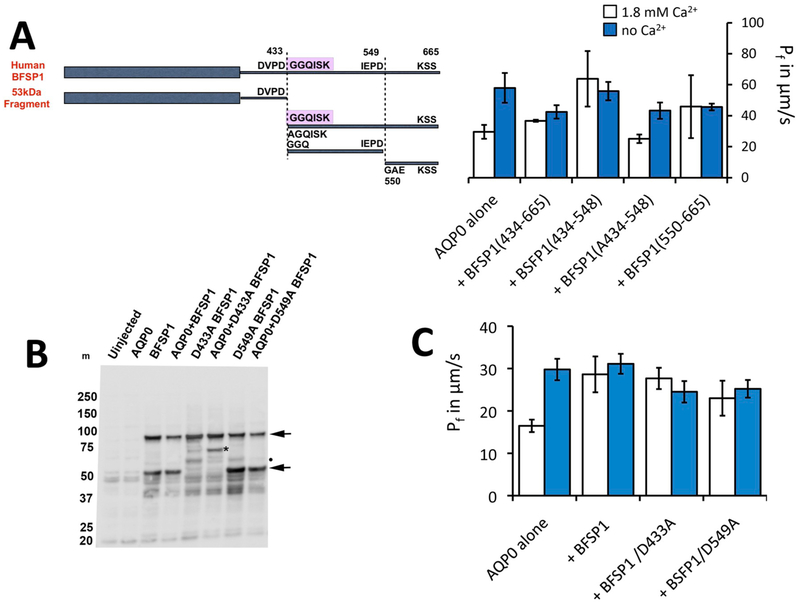
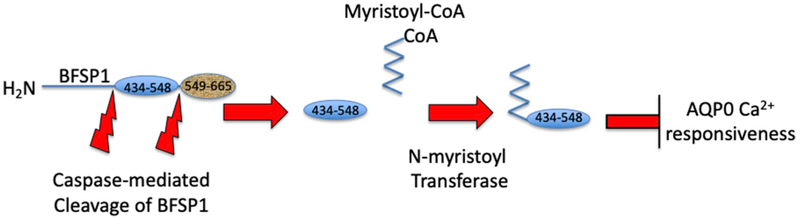
BFSP1 (beaded filament structural protein 1, filensin) is a cytoskeletal protein expressed in the eye lens. It binds AQP0 in vitro and its C-terminal sequences have been suggested to regulate the water channel activity of AQP0. A myristoylated fragment from the C-terminus of BFSP1 was found in AQP0 enriched fractions. Here we identify BFSP1 as a substrate for caspase-mediated cleavage at several C-terminal sites including D433. Cleavage at D433 exposes a cryptic myristoylation sequence (434-440). We confirm that this sequence is an excellent substrate for both NMT1 and 2 (N-myristoyl transferase). Thus caspase cleavage may promote formation of myristoylated fragments derived from the BFSP1 C-terminus (G434-S665). Myristoylation at G434 is not required for membrane association. Biochemical fractionation and immunogold labeling confirmed that C-terminal BFSP1 fragments containing the myristoylation sequence colocalized with AQP0 in the same plasma membrane compartments of lens fibre cells. To determine the functional significance of the association of BFSP1 G434-S665 sequences with AQP0, we measured AQP0 water permeability in Xenopus oocytes co-transfected with transcripts expressing both AQP0 and various C-terminal domain fragments of BFSP1 generated by caspase cleavage. We found that different fragments dramatically alter the response of AQP0 to different concentrations of Ca2+. The complete C-terminal fragment (G434-S665) eliminates calcium regulation altogether. Shorter fragments can enhance regulation by elevated calcium or reverse the response, indicative of the regulatory potential of BFSP1 with respect to AQP0. In particular, elimination of the myristoylation site by the mutation G434A reverses the order of water permeability sensitivity to different Ca2+ concentrations.
Copyright © 2019 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures
References
-
- Bologna G, Yvon C, Duvaud S, Veuthey AL, 2004. N-Terminal myristoylation predictions by ensembles of neural networks. Proteomics 4, 1626–1632. - PubMed
-
- Brunkener M, Georgatos SD, 1992. Membrane-binding properties of filensin, a cytoskeletal protein of the lens fiber cells. J. Cell Sci 103 (Pt 3), 709–718. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
