

Regulation of leukocyte function by citric acid cycle intermediates
- PMID: 30791134
- PMCID: PMC6597293
- DOI: 10.1002/JLB.3MIR1118-415R
Regulation of leukocyte function by citric acid cycle intermediates
Abstract
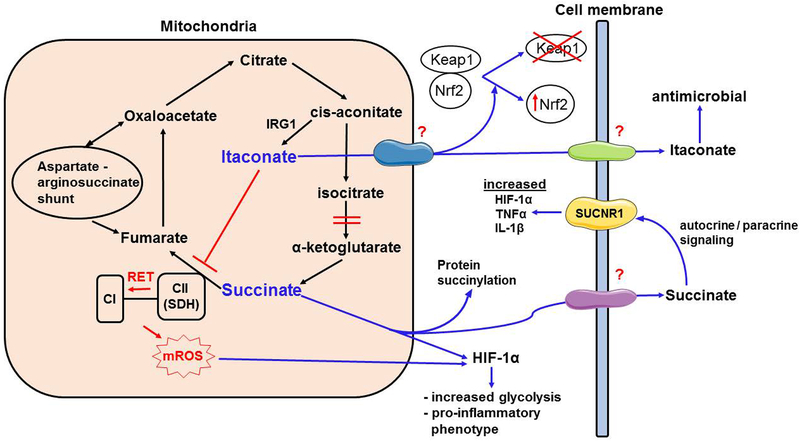
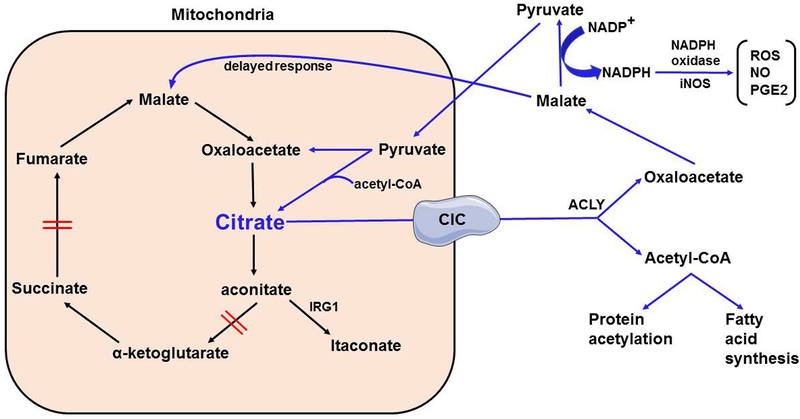
Cellular metabolism is a means of generating ATP to provide energy for key cellular functions. However, recent research shows that citric acid cycle intermediates target vital cellular functions of the innate immune system. Succinate, itaconate, citrate, and fumarate have been shown to mediate or regulate important myeloid cell functions during infection and inflammation. This review covers the regulatory functions of citric acid cycle intermediates in myeloid cells and discusses potential translational applications, key mechanistic questions, and future research directions.
Keywords: citrate; fumarate; itaconate; metabolic reprogramming; succinate.
©2019 Society for Leukocyte Biology.
Conflict of interest statement
Disclosure of Conflicts of Interest
Authors declare no conflict of interest.
Figures
Similar articles
-
Gang of 3: How the Krebs cycle-linked metabolites itaconate, succinate, and fumarate regulate macrophages and inflammation.Cell Metab. 2025 May 6;37(5):1049-1059. doi: 10.1016/j.cmet.2025.03.004. Epub 2025 Mar 31. Cell Metab. 2025. PMID: 40169002 Review.
-
Krebs Cycle Reimagined: The Emerging Roles of Succinate and Itaconate as Signal Transducers.Cell. 2018 Aug 9;174(4):780-784. doi: 10.1016/j.cell.2018.07.030. Cell. 2018. PMID: 30096309 Review.
-
Effect of citric acid-cycle intermediates on oxaloacetate utilization and succinate oxidation.Biochem J. 1960 Aug;76(2):293-7. doi: 10.1042/bj0760293. Biochem J. 1960. PMID: 13840087 Free PMC article. No abstract available.
-
THE UTILIZATION OF ACONATE AND ITACONATE BY MICROCOCCUS SP.Biochem J. 1965 Jan;94(1):25-31. doi: 10.1042/bj0940025. Biochem J. 1965. PMID: 14342240 Free PMC article.
-
Citric acid cycle intermediates as ligands for orphan G-protein-coupled receptors.Nature. 2004 May 13;429(6988):188-93. doi: 10.1038/nature02488. Nature. 2004. PMID: 15141213
Cited by
-
TLR Agonists as Mediators of Trained Immunity: Mechanistic Insight and Immunotherapeutic Potential to Combat Infection.Front Immunol. 2021 Feb 18;11:622614. doi: 10.3389/fimmu.2020.622614. eCollection 2020. Front Immunol. 2021. PMID: 33679711 Free PMC article. Review.
-
Tricarboxylic Acid (TCA) Cycle Intermediates: Regulators of Immune Responses.Life (Basel). 2021 Jan 19;11(1):69. doi: 10.3390/life11010069. Life (Basel). 2021. PMID: 33477822 Free PMC article. Review.
-
Metabolic insights into HIV/TB co-infection: an untargeted urinary metabolomics approach.Metabolomics. 2024 Jul 16;20(4):78. doi: 10.1007/s11306-024-02148-5. Metabolomics. 2024. PMID: 39014031 Free PMC article.
-
Mitochondrial citrate accumulation drives alveolar epithelial cell necroptosis in lipopolysaccharide-induced acute lung injury.Exp Mol Med. 2022 Nov;54(11):2077-2091. doi: 10.1038/s12276-022-00889-8. Epub 2022 Nov 28. Exp Mol Med. 2022. PMID: 36443565 Free PMC article.
-
Immunoresponsive gene 1 facilitates TLR4 agonist-induced augmentation of innate antimicrobial immunity.J Leukoc Biol. 2025 Feb 13;117(2):qiae198. doi: 10.1093/jleuko/qiae198. J Leukoc Biol. 2025. PMID: 39351765 Free PMC article.
References
-
- Tannahill GM, Curtis AM, Adamik J, Palsson-McDermott EM, McGettrick AF, Goel G, Frezza C, Bernard NJ, Kelly B, Foley NH, Zheng L, Gardet A, Tong Z, Jany SS, Corr SC, Haneklaus M, Caffrey BE, Pierce K, Walmsley S, Beasley FC, Cummins E, Nizet V, Whyte M, Taylor CT, Lin H, Masters SL, Gottlieb E, Kelly VP, Clish C, Auron PE, Xavier RJ, O’Neill LA (2013) Succinate is an inflammatory signal that induces IL-1beta through HIF-1alpha. Nature 496, 238–42. - PMC - PubMed
-
- Lampropoulou V, Sergushichev A, Bambouskova M, Nair S, Vincent EE, Loginicheva E, Cervantes-Barragan L, Ma X, Huang SC, Griss T, Weinheimer CJ, Khader S, Randolph GJ, Pearce EJ, Jones RG, Diwan A, Diamond MS, Artyomov MN (2016) Itaconate Links Inhibition of Succinate Dehydrogenase with Macrophage Metabolic Remodeling and Regulation of Inflammation. Cell Metab 24, 158–66. - PMC - PubMed
-
- Murphy MP and O’Neill LAJ (2018) Krebs Cycle Reimagined: The Emerging Roles of Succinate and Itaconate as Signal Transducers. Cell 174, 780–784. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
