

An NAD+ Phosphorylase Toxin Triggers Mycobacterium tuberculosis Cell Death
- PMID: 30792174
- PMCID: PMC6436930
- DOI: 10.1016/j.molcel.2019.01.028
An NAD+ Phosphorylase Toxin Triggers Mycobacterium tuberculosis Cell Death
Abstract
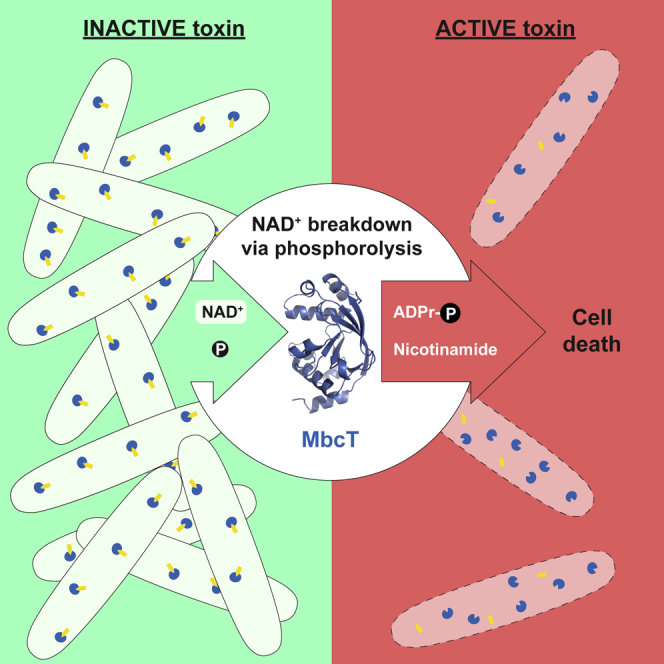
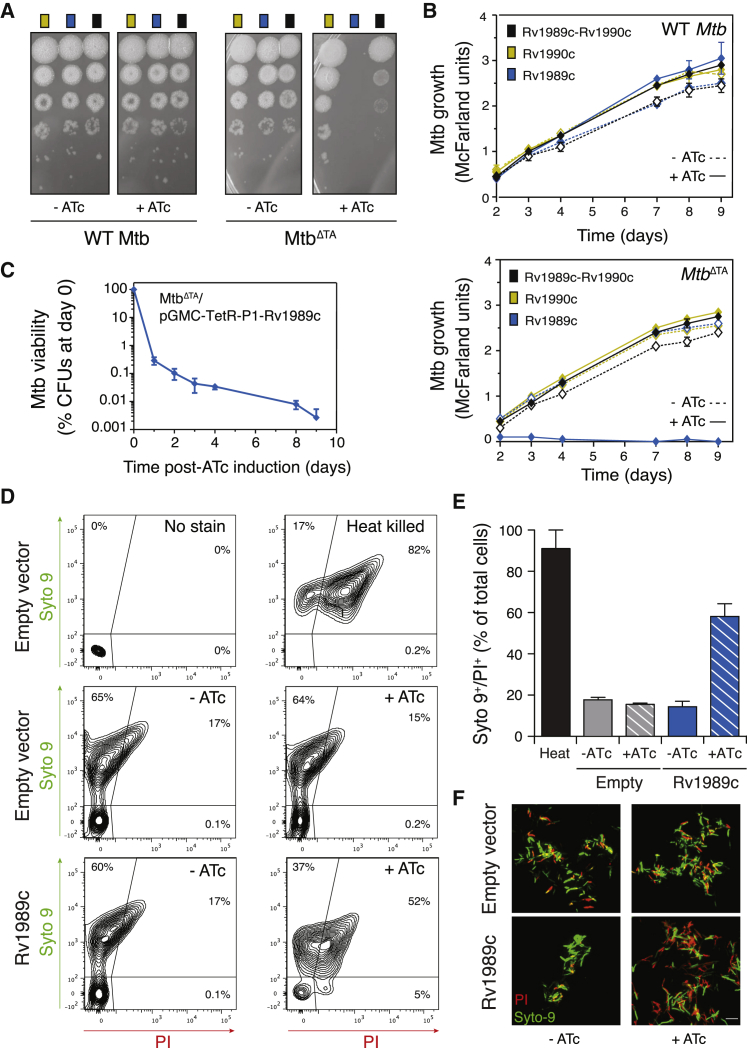
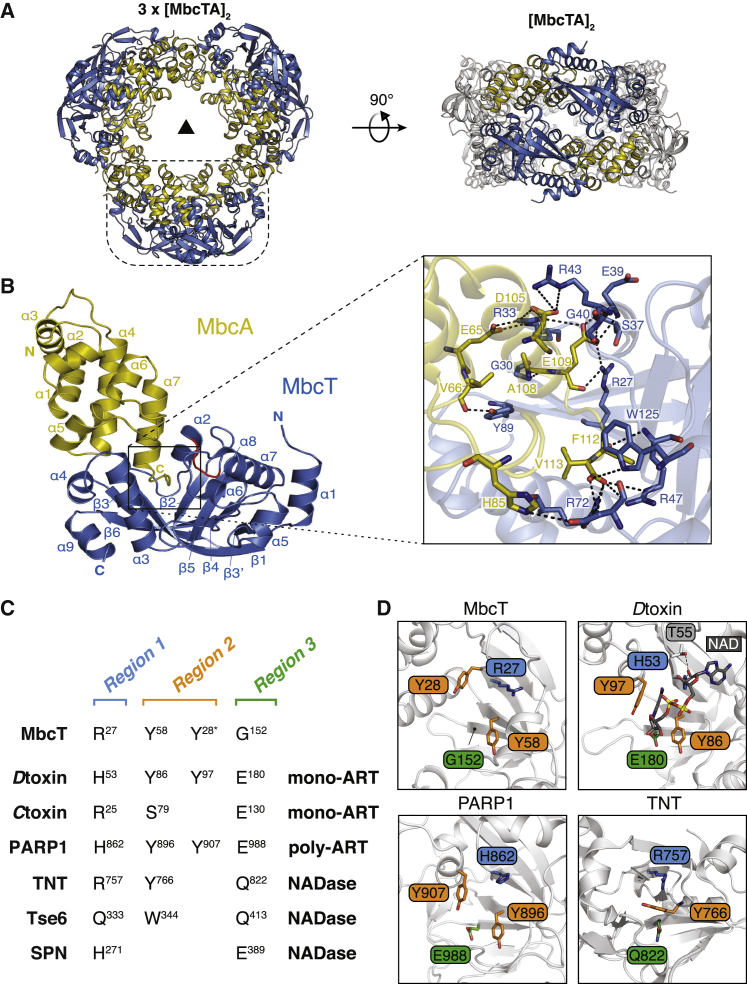
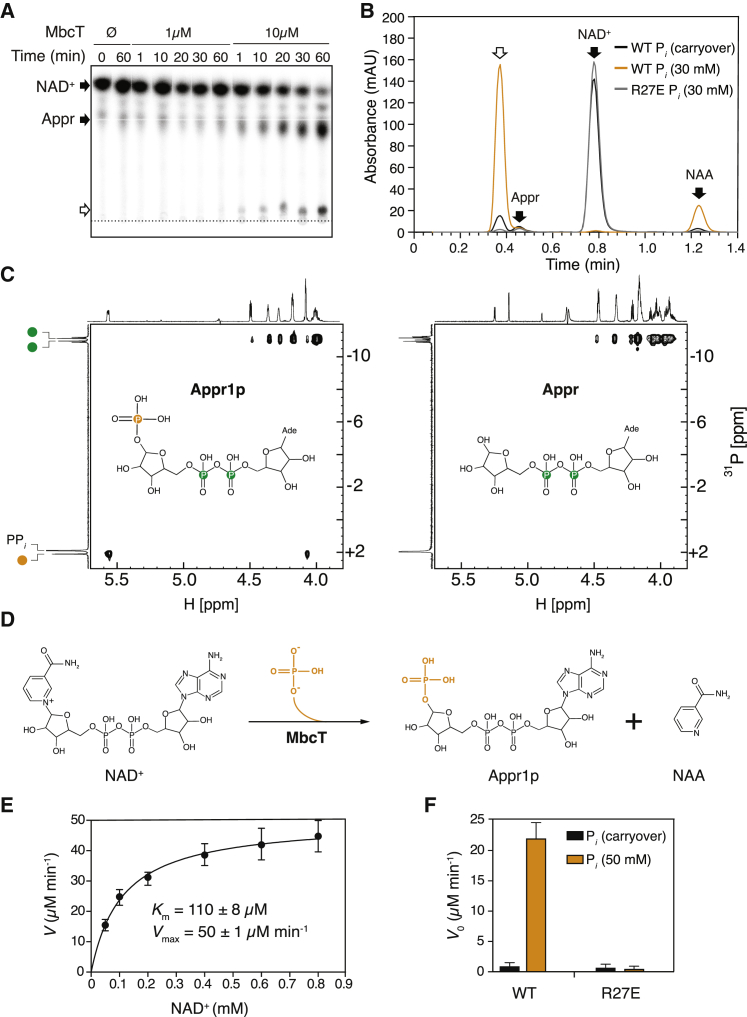
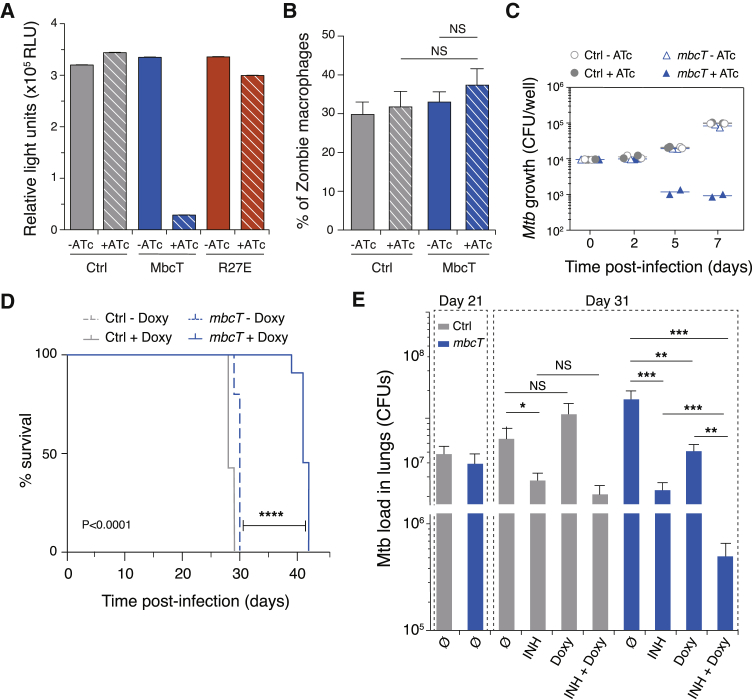
Toxin-antitoxin (TA) systems regulate fundamental cellular processes in bacteria and represent potential therapeutic targets. We report a new RES-Xre TA system in multiple human pathogens, including Mycobacterium tuberculosis. The toxin, MbcT, is bactericidal unless neutralized by its antitoxin MbcA. To investigate the mechanism, we solved the 1.8 Å-resolution crystal structure of the MbcTA complex. We found that MbcT resembles secreted NAD+-dependent bacterial exotoxins, such as diphtheria toxin. Indeed, MbcT catalyzes NAD+ degradation in vitro and in vivo. Unexpectedly, the reaction is stimulated by inorganic phosphate, and our data reveal that MbcT is a NAD+ phosphorylase. In the absence of MbcA, MbcT triggers rapid M. tuberculosis cell death, which reduces mycobacterial survival in macrophages and prolongs the survival of infected mice. Our study expands the molecular activities employed by bacterial TA modules and uncovers a new class of enzymes that could be exploited to treat tuberculosis and other infectious diseases.
Keywords: MbcTA; NAD; bacterial cell death; mycobacterium; toxin-antitoxin system; tuberculosis.
Crown Copyright © 2019. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Becq J., Gutierrez M.C., Rosas-Magallanes V., Rauzier J., Gicquel B., Neyrolles O., Deschavanne P. Contribution of horizontally acquired genomic islands to the evolution of the Tubercle bacilli. Mol. Biol. Evol. 2007;24:1861–1871. - PubMed
-
- Blanchet C.E., Spilotros A., Schwemmer F., Graewert M.A., Kikhney A., Jeffries C.M., Franke D., Mark D., Zengerle R., Cipriani F. Versatile sample environments and automation for biological solution X-ray scattering experiments at the P12 beamline (PETRA III, DESY) J. Appl. Cryst. 2015;48:431–443. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
