

Autophagic flux modulation by Wnt/β-catenin pathway inhibition in hepatocellular carcinoma
- PMID: 30794613
- PMCID: PMC6386480
- DOI: 10.1371/journal.pone.0212538
Autophagic flux modulation by Wnt/β-catenin pathway inhibition in hepatocellular carcinoma
Abstract
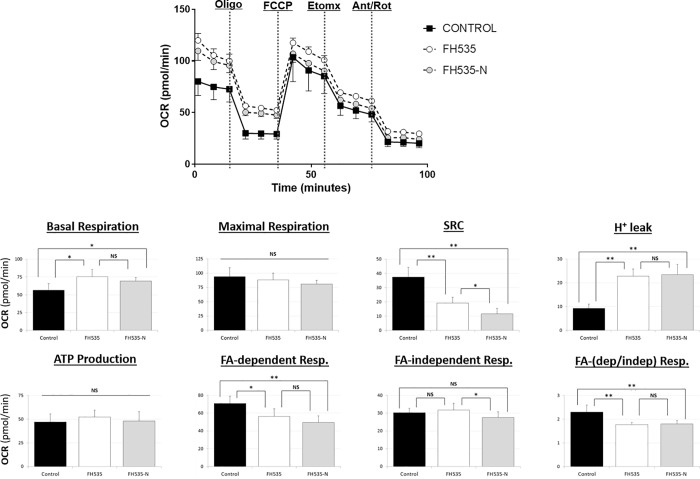
Autophagy targets cellular components for lysosomal-dependent degradation in which the products of degradation may be recycled for protein synthesis and utilized for energy production. Autophagy also plays a critical role in cell homeostasis and the regulation of many physiological and pathological processes and prompts this investigation of new agents to effect abnormal autophagy in hepatocellular carcinoma (HCC). 2,5-Dichloro-N-(2-methyl-4-nitrophenyl) benzenesulfonamide (FH535) is a synthetic inhibitor of the Wnt/β-catenin pathway that exhibits anti-proliferative and anti-angiogenic effects on different types of cancer cells. The combination of FH535 with sorafenib promotes a synergistic inhibition of HCC and liver cancer stem cell proliferation, mediated in part by the simultaneous disruption of mitochondrial respiration and glycolysis. We demonstrated that FH535 decreased HCC tumor progression in a mouse xenograft model. For the first time, we showed the inhibitory effect of an FH535 derivative, FH535-N, alone and in combination with sorafenib on HCC cell proliferation. Our study revealed the contributing effect of Wnt/β-catenin pathway inhibition by FH535 and its derivative (FH535-N) through disruption of the autophagic flux in HCC cells.
Conflict of interest statement
CL and DSW have partial ownership in a private venture, Epionc, Inc., incorporated to develop small-molecule inhibitors for cancer treatment through a licensing agreement with the University of Kentucky. In accord with University of Kentucky policies, CL and DSW disclosed prior work to the University of Kentucky’s Intellectual Property Committee and complied with stipulations of the University’s Conflict of Interest Oversight Committee. Epionc, Inc., has no license for compounds described in this paper. This does not alter our adherence to PLOS ONE policies on sharing data and materials.
Figures










References
-
- Society AC. Facts & Figures 2018 Atlanta, Ga.2018 [cited 2018 Sept 9]. Available from: https://www.cancer.org/cancer/liver-cancer/about/what-is-key-statistics.....
-
- Altekruse SF, McGlynn KA, Reichman ME. Hepatocellular carcinoma incidence, mortality, and survival trends in the United States from 1975 to 2005. Journal of clinical oncology: official journal of the American Society of Clinical Oncology. 2009;27(9):1485–91. Epub 2009/02/20. 10.1200/jco.2008.20.7753 - DOI - PMC - PubMed
-
- Dimitroulis D, Damaskos C, Valsami S, Davakis S, Garmpis N, Spartalis E, et al. From diagnosis to treatment of hepatocellular carcinoma: An epidemic problem for both developed and developing world. World journal of gastroenterology: WJG. 2017;23(29):5282–94. Epub 2017/08/26. 10.3748/wjg.v23.i29.5282 - DOI - PMC - PubMed
-
- Lee HC, Kim M, Wands JR. Wnt/Frizzled signaling in hepatocellular carcinoma. Frontiers in bioscience: a journal and virtual library. 2006;11:1901–15. Epub 2005/12/22. . - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
