

Mathematical Modelling of Endocrine Systems
- PMID: 30799185
- PMCID: PMC6425086
- DOI: 10.1016/j.tem.2019.01.008
Mathematical Modelling of Endocrine Systems
Abstract
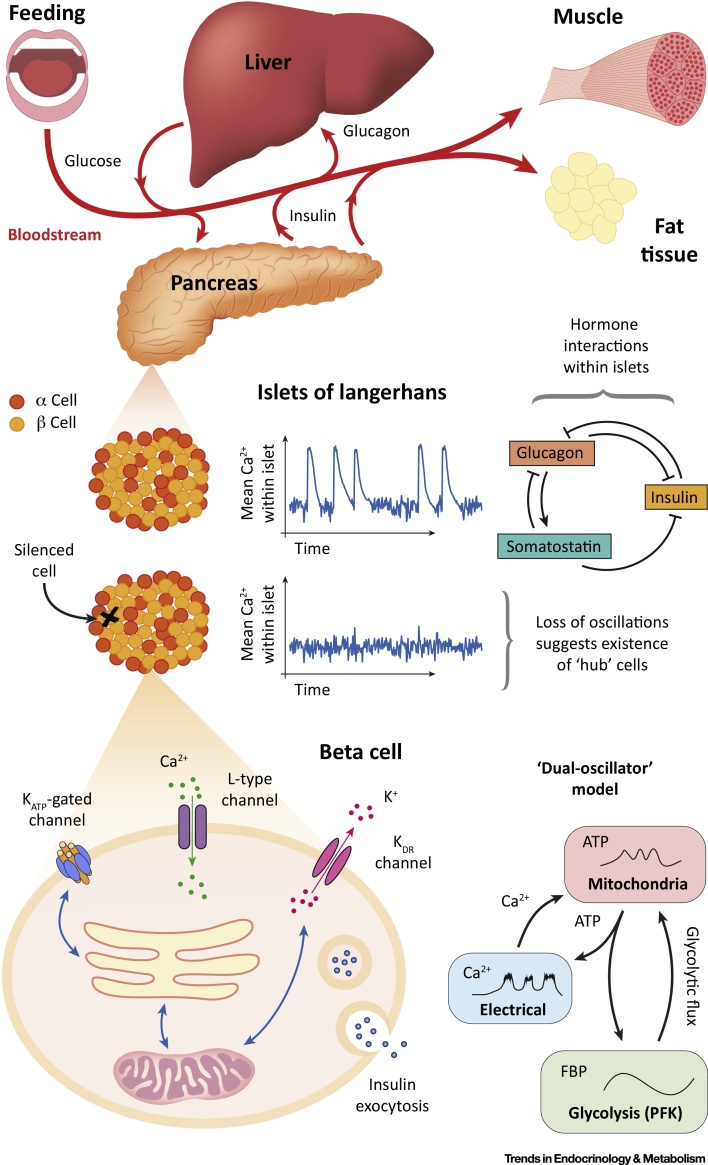
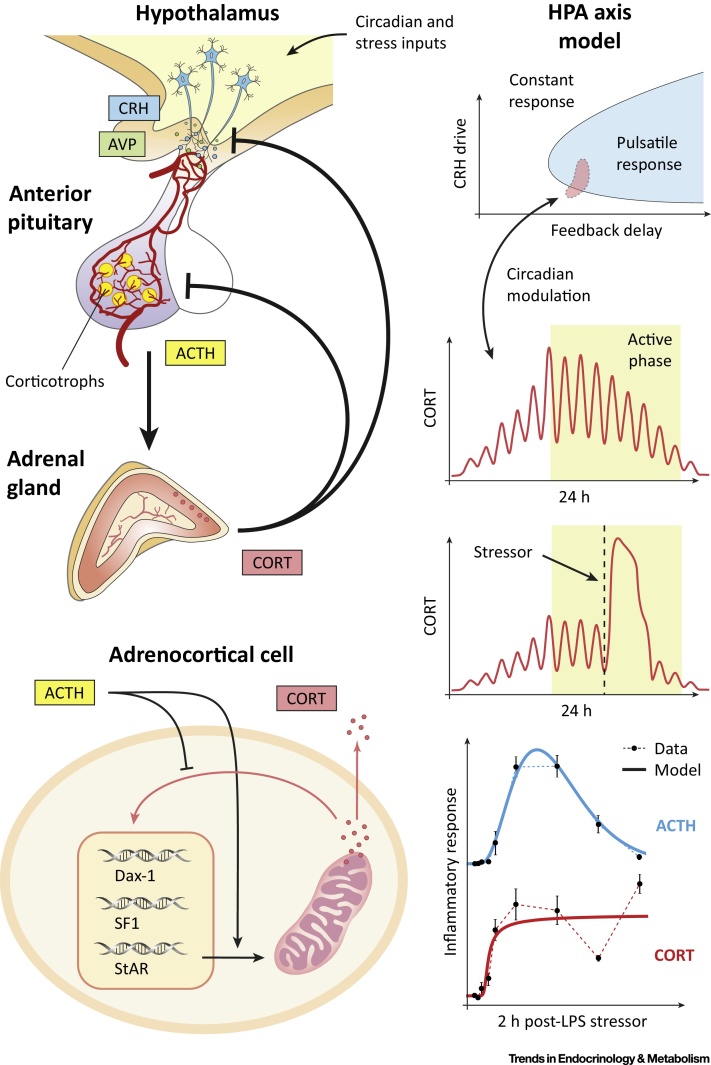
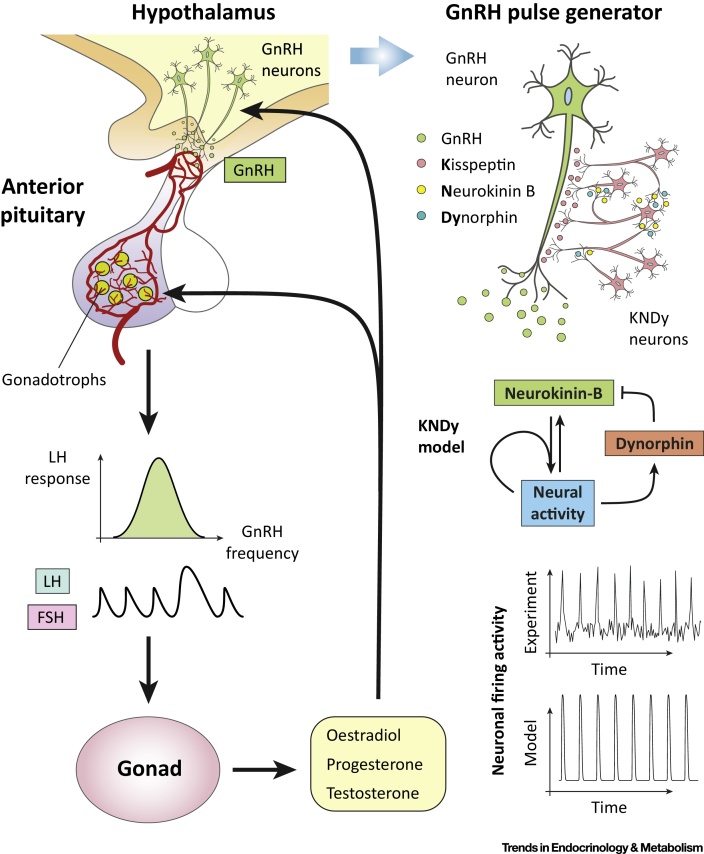
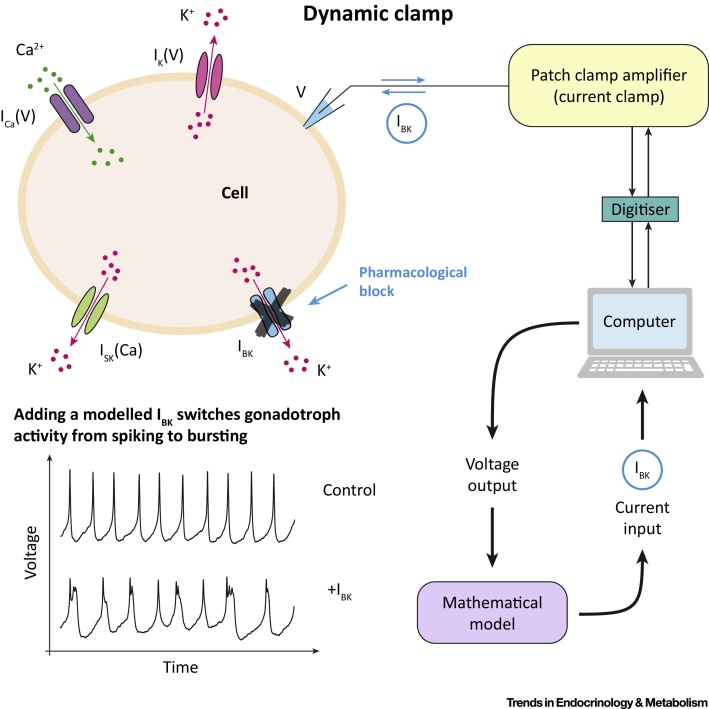
Hormone rhythms are ubiquitous and essential to sustain normal physiological functions. Combined mathematical modelling and experimental approaches have shown that these rhythms result from regulatory processes occurring at multiple levels of organisation and require continuous dynamic equilibration, particularly in response to stimuli. We review how such an interdisciplinary approach has been successfully applied to unravel complex regulatory mechanisms in the metabolic, stress, and reproductive axes. We discuss how this strategy is likely to be instrumental for making progress in emerging areas such as chronobiology and network physiology. Ultimately, we envisage that the insight provided by mathematical models could lead to novel experimental tools able to continuously adapt parameters to gradual physiological changes and the design of clinical interventions to restore normal endocrine function.
Keywords: chronotherapy; circadian rhythms; hormone dynamics; hybrid systems; regulatory networks; ultradian oscillations.
Copyright © 2019 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures
References
-
- Leng G., Macgregor D.J. Mathematical modelling in neuroendocrinology. J. Neuroendocrinol. 2008;20:713–718. - PubMed
-
- Leng G., MacGregor D.J. Models in neuroendocrinology. Math. Biosci. 2018;305:29–41. - PubMed
-
- Bertram R. Mathematical modeling in neuroendocrinology. Compr. Physiol. 2015;5:911–927. - PubMed
-
- Nyman E. Insulin signaling – mathematical modeling comes of age. Trends Endocrinol. Metab. 2012;23:107–115. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
