

Cellular entry and uncoating of naked and quasi-enveloped human hepatoviruses
- PMID: 30801249
- PMCID: PMC6422491
- DOI: 10.7554/eLife.43983
Cellular entry and uncoating of naked and quasi-enveloped human hepatoviruses
Abstract
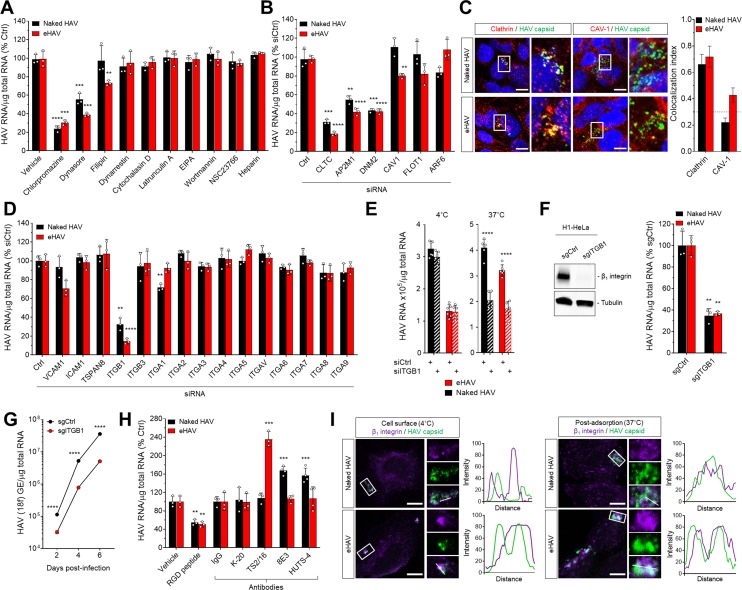
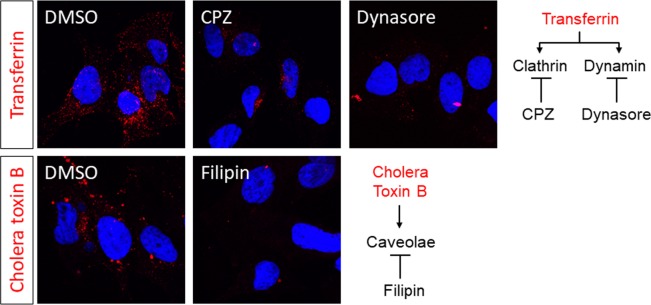
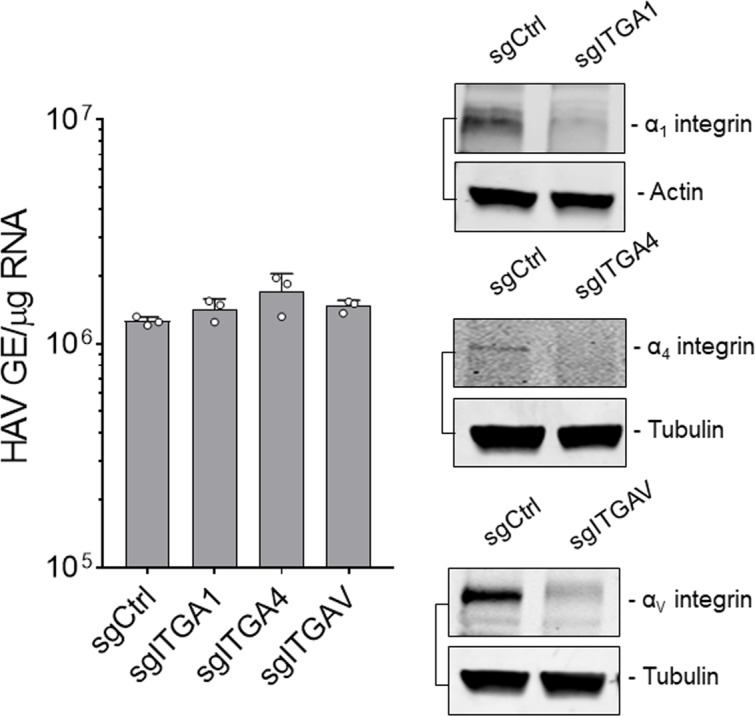
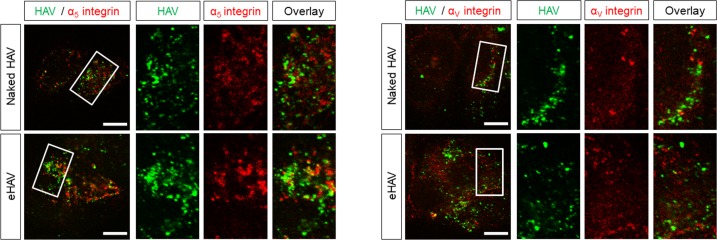
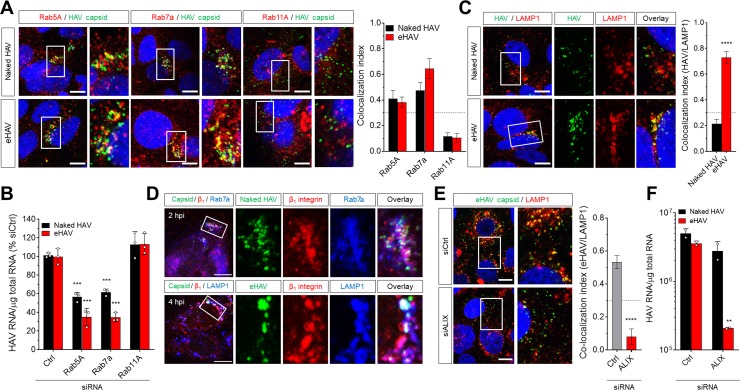
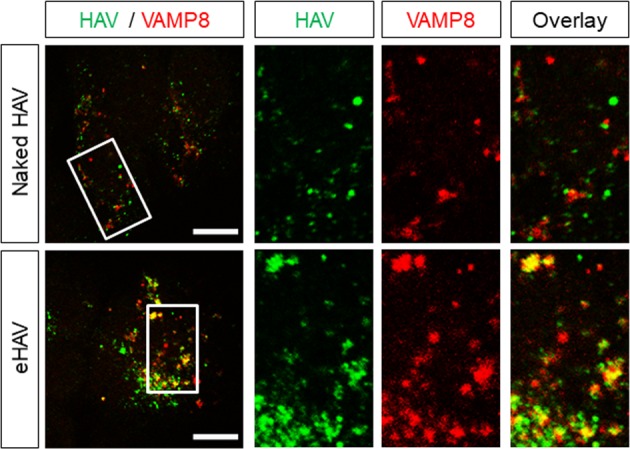
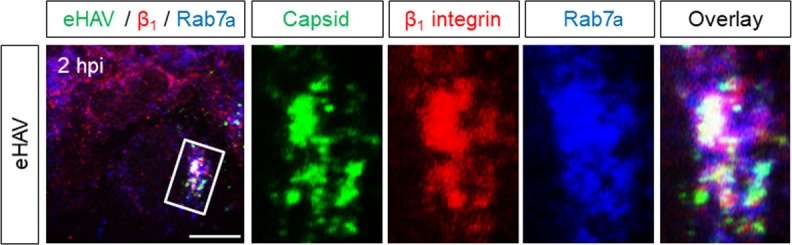
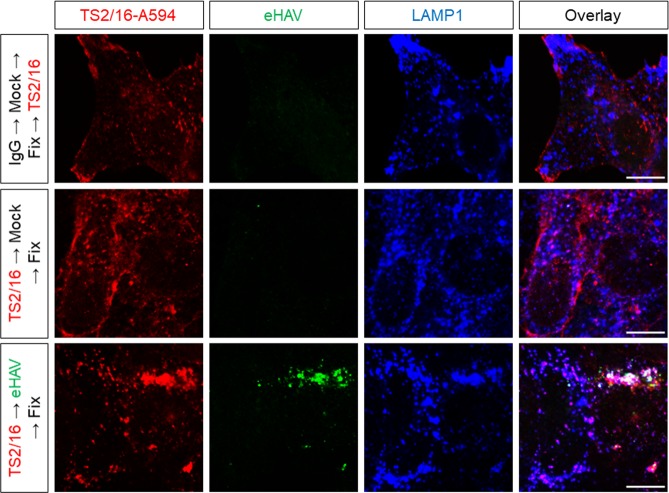
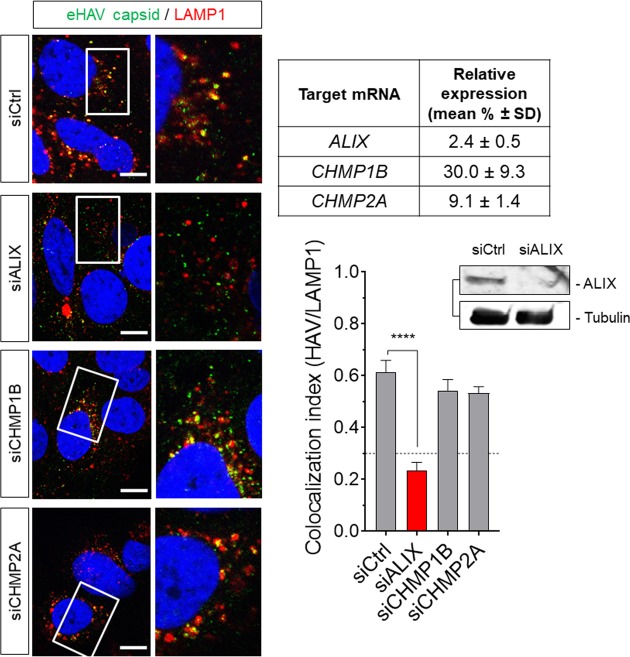
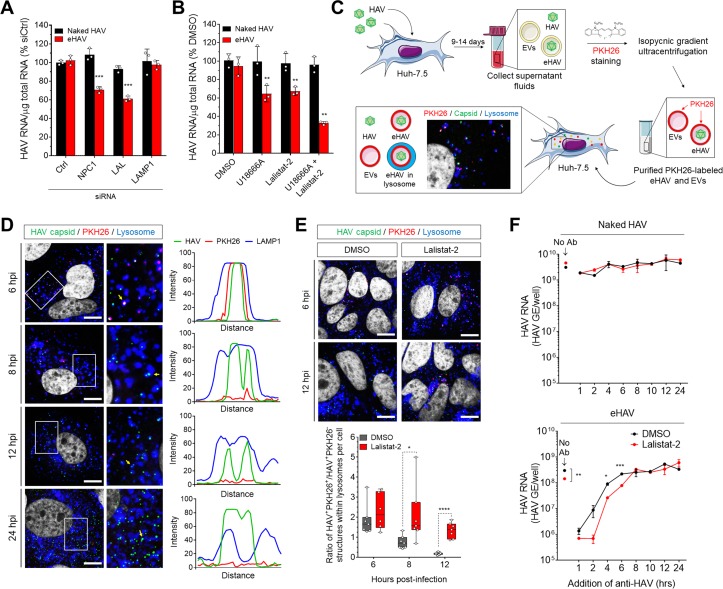
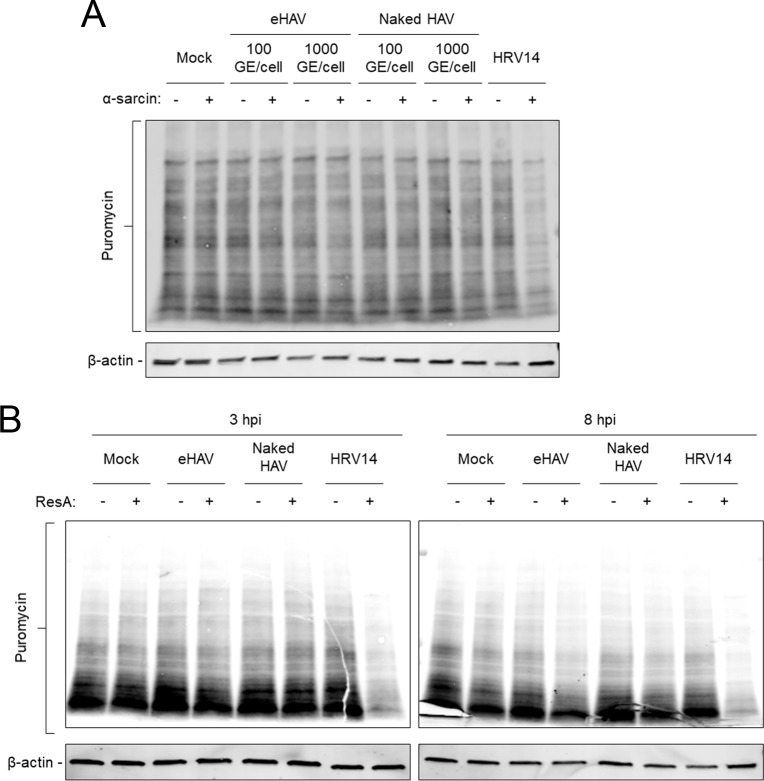
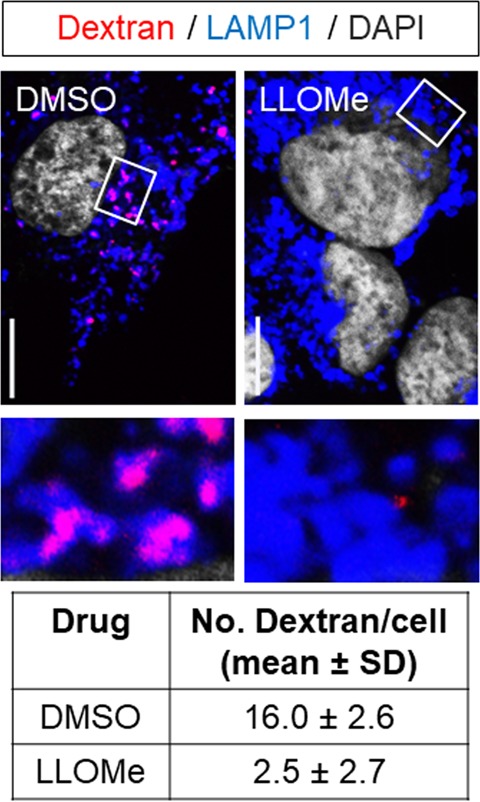
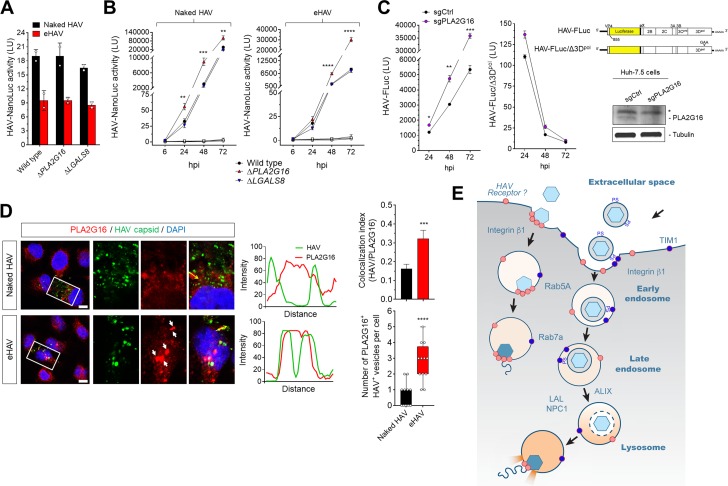
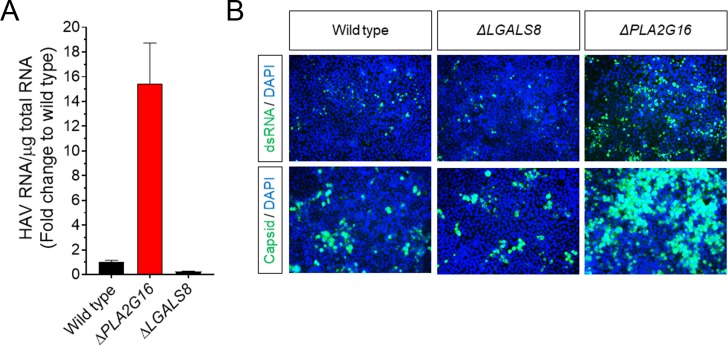
Many 'non-enveloped' viruses, including hepatitis A virus (HAV), are released non-lytically from infected cells as infectious, quasi-enveloped virions cloaked in host membranes. Quasi-enveloped HAV (eHAV) mediates stealthy cell-to-cell spread within the liver, whereas stable naked virions shed in feces are optimized for environmental transmission. eHAV lacks virus-encoded surface proteins, and how it enters cells is unknown. We show both virion types enter by clathrin- and dynamin-dependent endocytosis, facilitated by integrin β1, and traffic through early and late endosomes. Uncoating of naked virions occurs in late endosomes, whereas eHAV undergoes ALIX-dependent trafficking to lysosomes where the quasi-envelope is enzymatically degraded and uncoating ensues coincident with breaching of endolysosomal membranes. Neither virion requires PLA2G16, a phospholipase essential for entry of other picornaviruses. Thus naked and quasi-enveloped virions enter via similar endocytic pathways, but uncoat in different compartments and release their genomes to the cytosol in a manner mechanistically distinct from other Picornaviridae.
Keywords: PLA2G16; endocytic trafficking; exosomes; extracellular vesicles; infectious disease; integrins; microbiology; picornavirus; virus.
© 2019, Rivera-Serrano et al.
Conflict of interest statement
ER, OG, AD, SL No competing interests declared
Figures
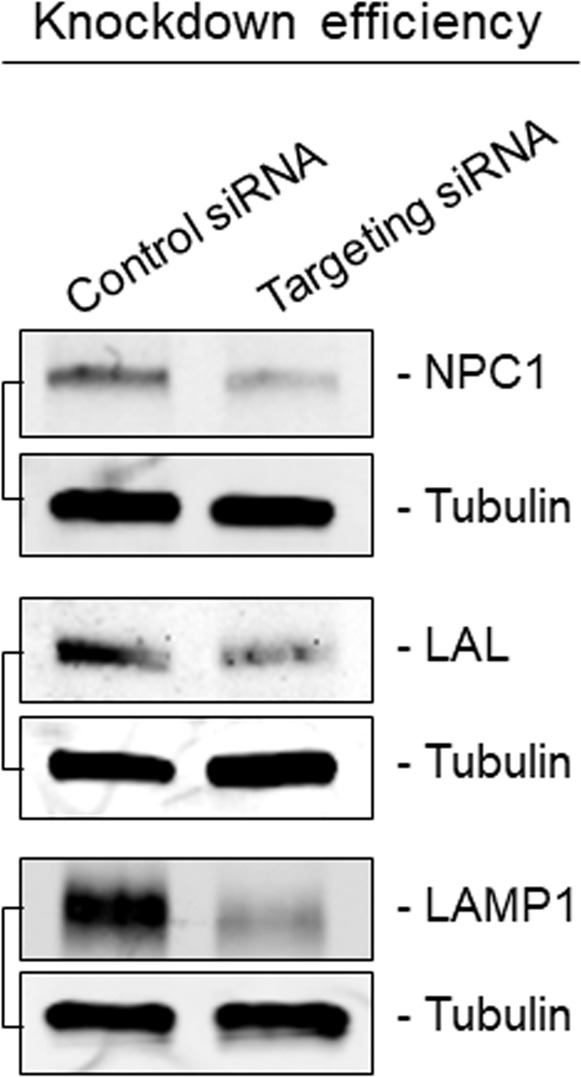
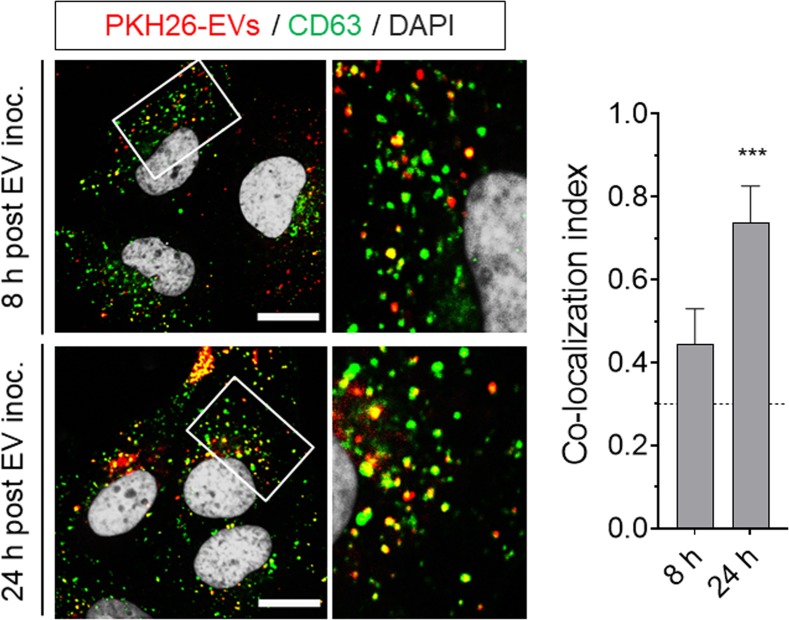
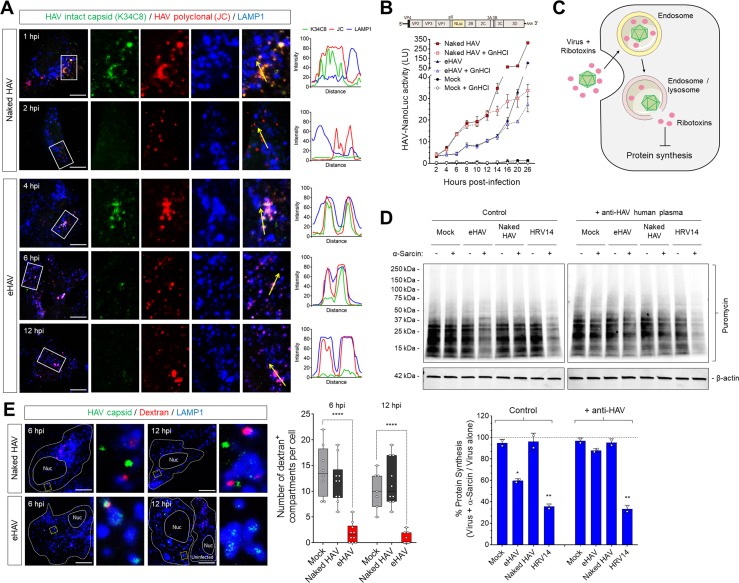
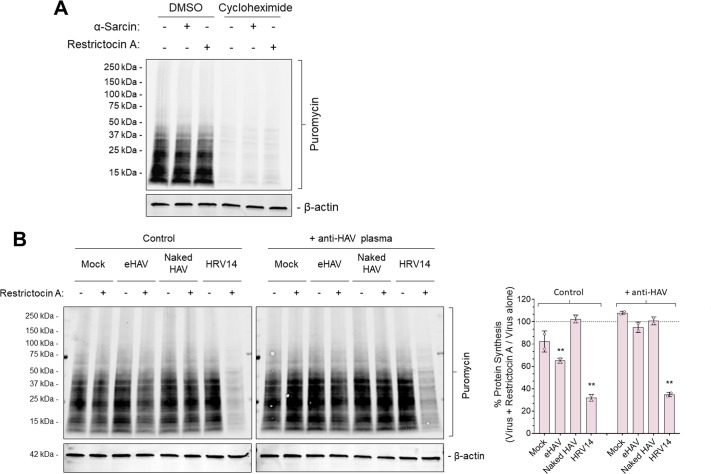
References
-
- Chen YH, Du W, Hagemeijer MC, Takvorian PM, Pau C, Cali A, Brantner CA, Stempinski ES, Connelly PS, Ma HC, Jiang P, Wimmer E, Altan-Bonnet G, Altan-Bonnet N. Phosphatidylserine vesicles enable efficient en bloc transmission of enteroviruses. Cell. 2015;160:619–630. doi: 10.1016/j.cell.2015.01.032. - DOI - PMC - PubMed
-
- Das A, Hirai-Yuki A, González-López O, Rhein B, Moller-Tank S, Brouillette R, Hensley L, Misumi I, Lovell W, Cullen JM, Whitmire JK, Maury W, Lemon SM. TIM1 (HAVCR1) Is not essential for cellular entry of either Quasi-enveloped or naked hepatitis A virions. mBio. 2017;8:e00969-17. doi: 10.1128/mBio.00969-17. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
