

Making and breaking heme
- PMID: 30802830
- PMCID: PMC6706330
- DOI: 10.1016/j.sbi.2019.01.006
Making and breaking heme
Abstract
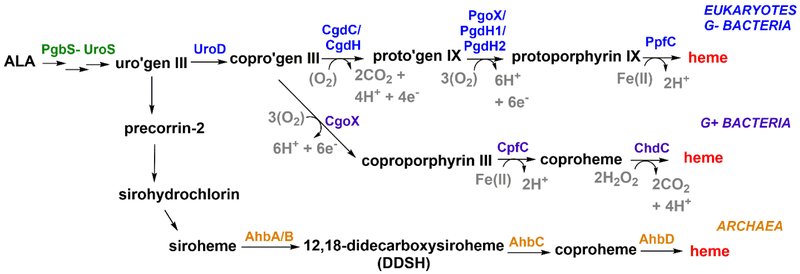
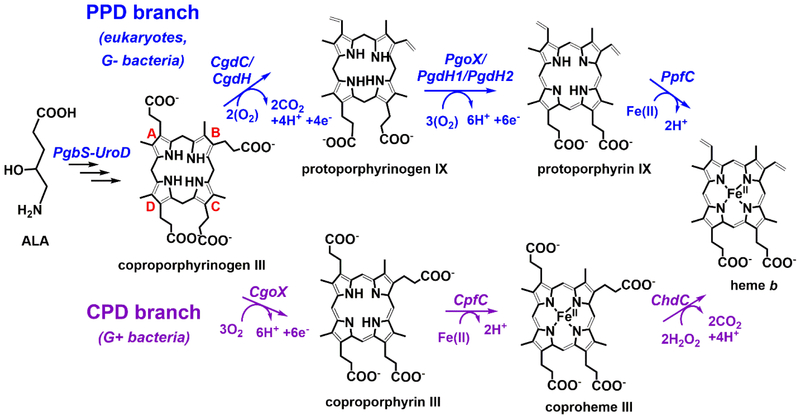
Mechanisms for making and breaking the heme b cofactor (heme) are more diverse than previously expected. Biosynthetic pathways have diverged at least twice along taxonomic lines, reflecting differences in membrane organization and O2 utilization among major groups of organisms. At least three families of heme degradases are now known, again differing in whether and how O2 is used by the organism and possibly the purpose for turning over the tetrapyrrole. Understanding these enzymes and pathways offers a handle for antimicrobial development and for monitoring heme use in organismal and ecological systems.
Copyright © 2019 Elsevier Ltd. All rights reserved.
Figures
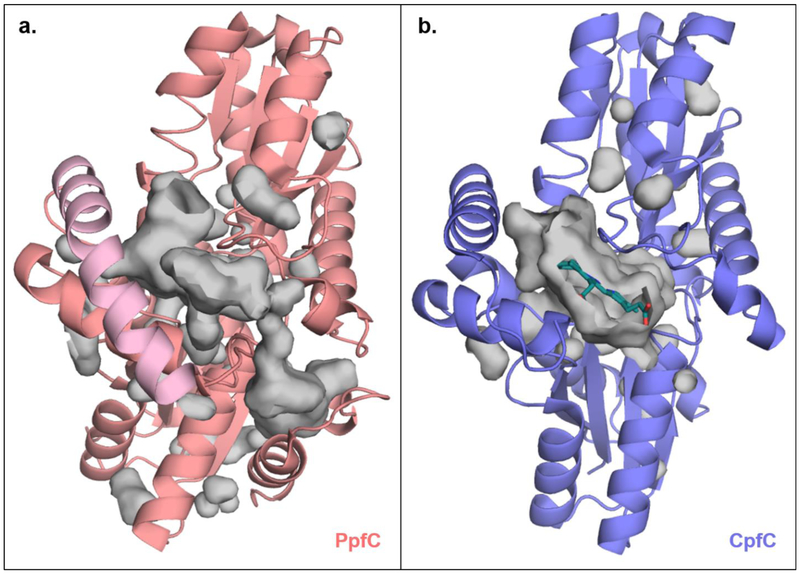
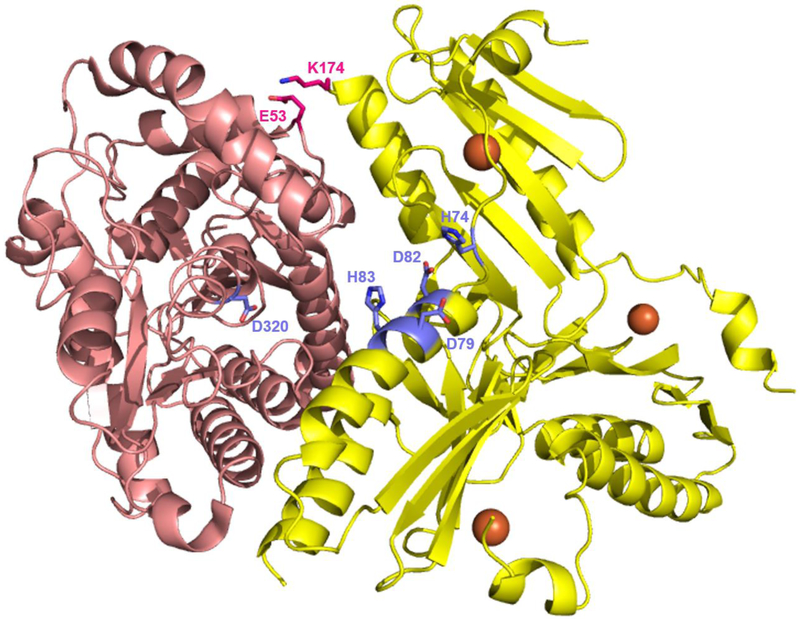
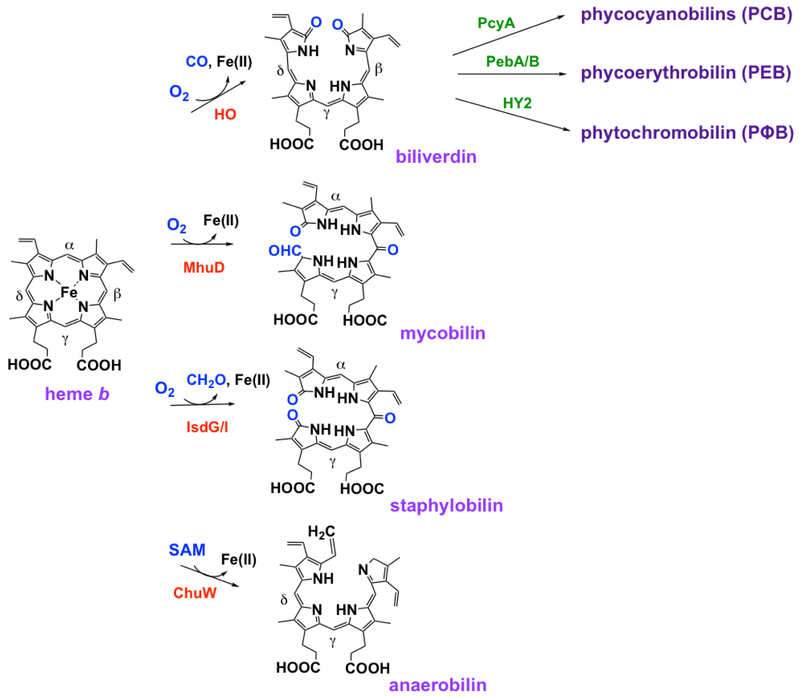
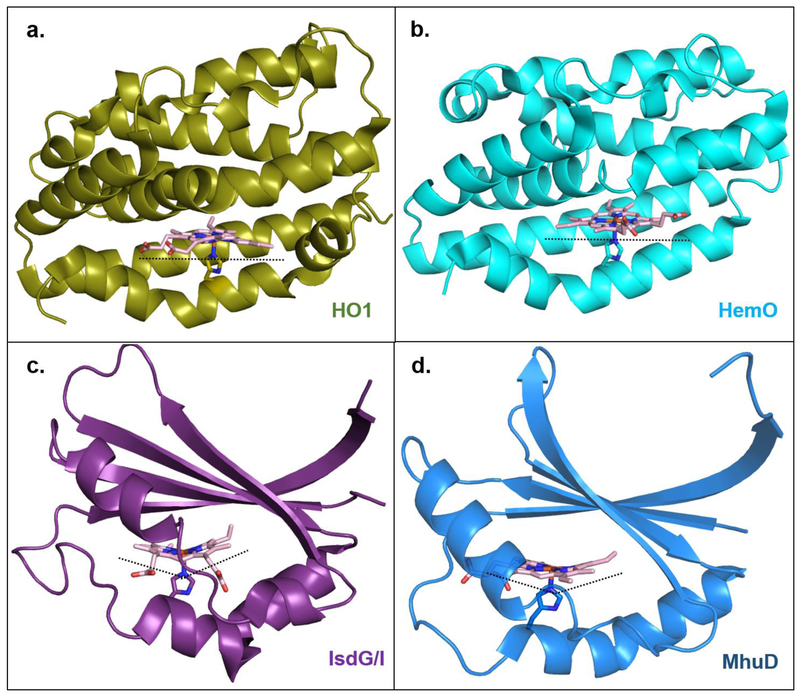
References
-
- Panek H, O'Brian MR: A whole genome view of prokaryotic haem biosynthesis. Microbiology 2002, 148:2273–2282. - PubMed
-
-
Bali S, Lawrence AD, Lobo SA, Saraiva LM, Golding BT, Palmer DJ, Howard MJ, Ferguson SJ, Warren MJ: Molecular hijacking of siroheme for the synthesis of heme and d1 heme. Proc Natl Acad Sci U S A 2011, 108:18260–18265.
• First article to describe a heme biosynthetic pathway for Archaea that differs from the canonical (PPD branch) heme biosynthesis pathway.
-
-
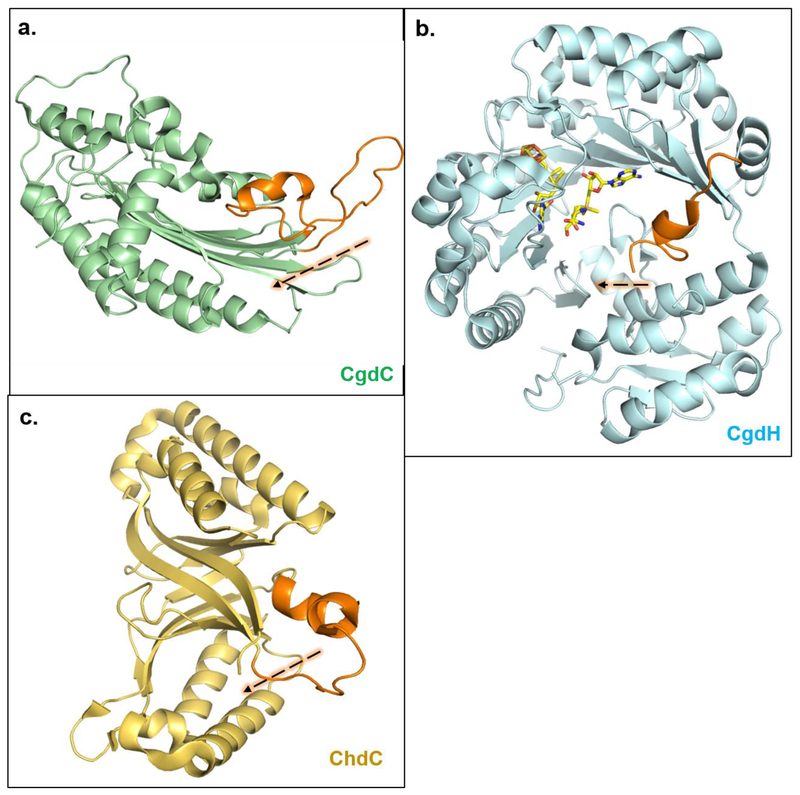
- Phillips JD, Whitby FG, Warby CA, Labbe P, Yang C, Pflugrath JW, Ferrara JD, Robinson H, Kushner JP, Hill CP: Crystal structure of the oxygen-dependant coproporphyrinogen oxidase (Hem13p) of Saccharomyces cerevisiae. Journal of Biological Chemistry 2004, 279:38960–38968. - PubMed
-
-
Dailey HA, Dailey TA, Gerdes S, Jahn D, Jahn M, O'Brian MR, Warren MJ: Prokaryotic Heme biosynthesis: Multiple pathways to a common essential product. Microbiol Mol Biol Rev 2017, 81.
•• Extensive and thorough review article that describes and compares all heme biosynthesis pathways, and the involved enzymes, known to date; the PPD branch, the CPD branch, and the Ahb pathway branch.
-
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
