

SHARPIN at the nexus of integrin, immune, and inflammatory signaling in human platelets
- PMID: 30804189
- PMCID: PMC6421413
- DOI: 10.1073/pnas.1819156116
SHARPIN at the nexus of integrin, immune, and inflammatory signaling in human platelets
Abstract
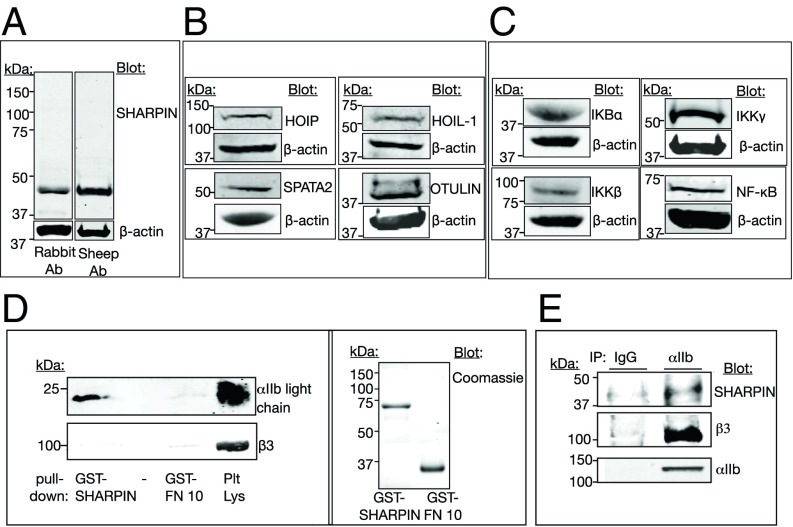
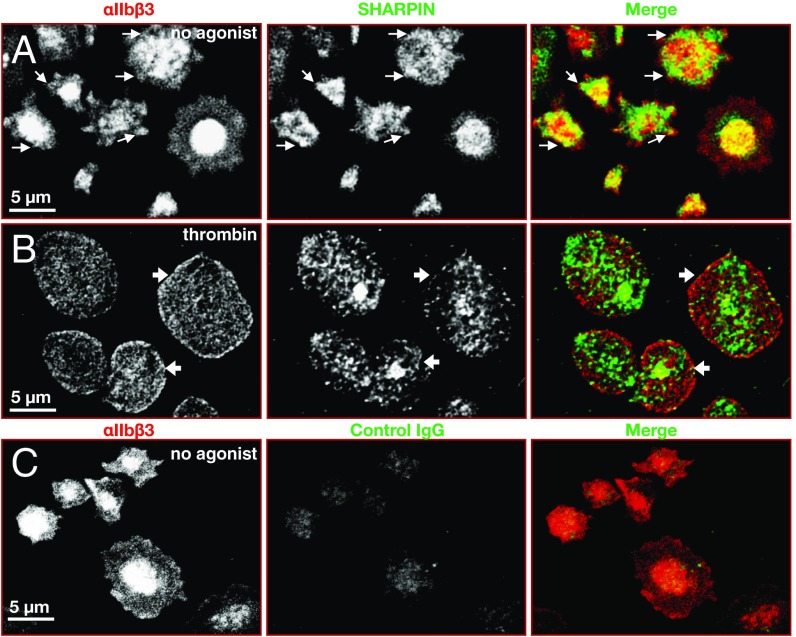
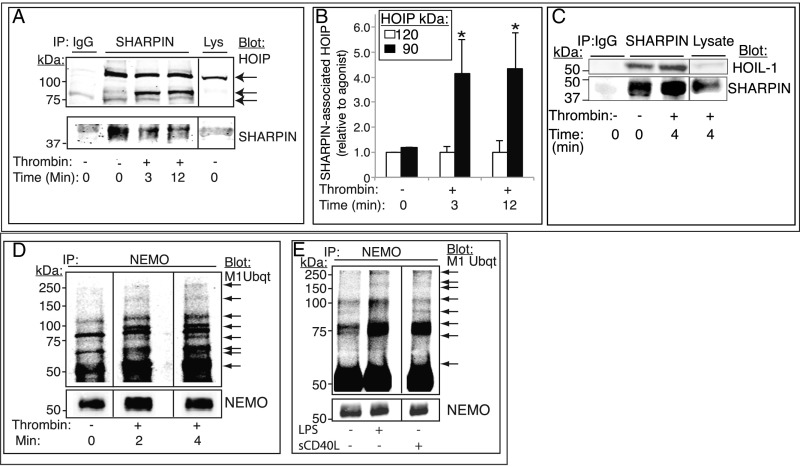
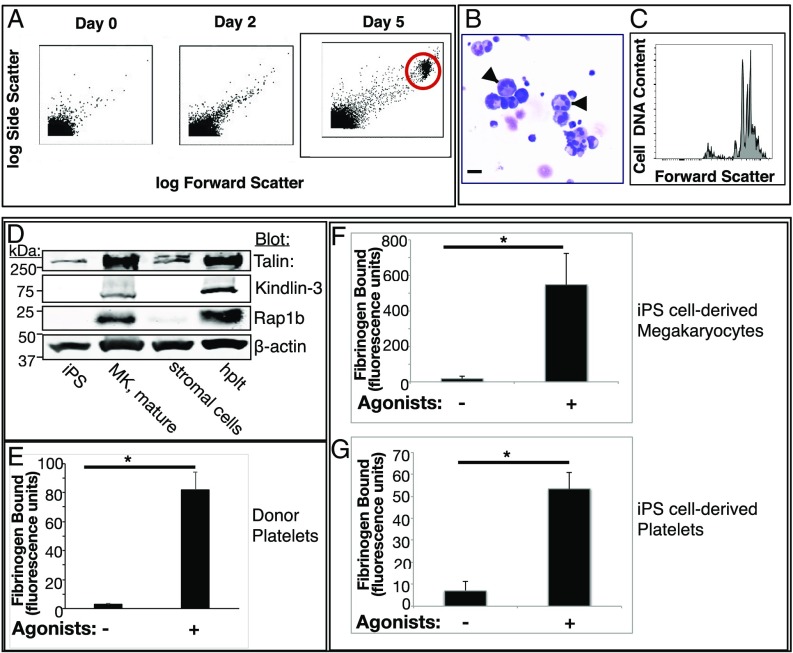
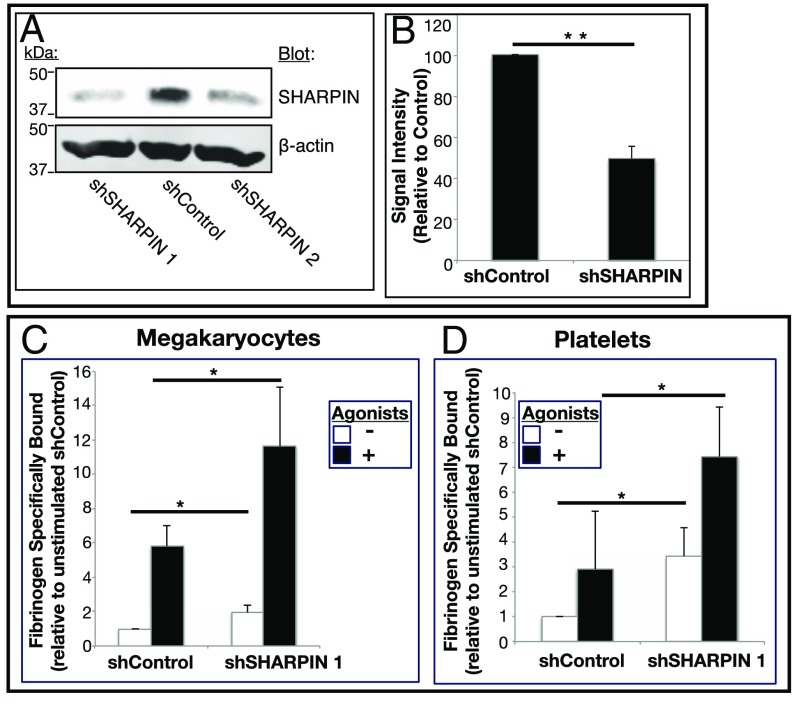
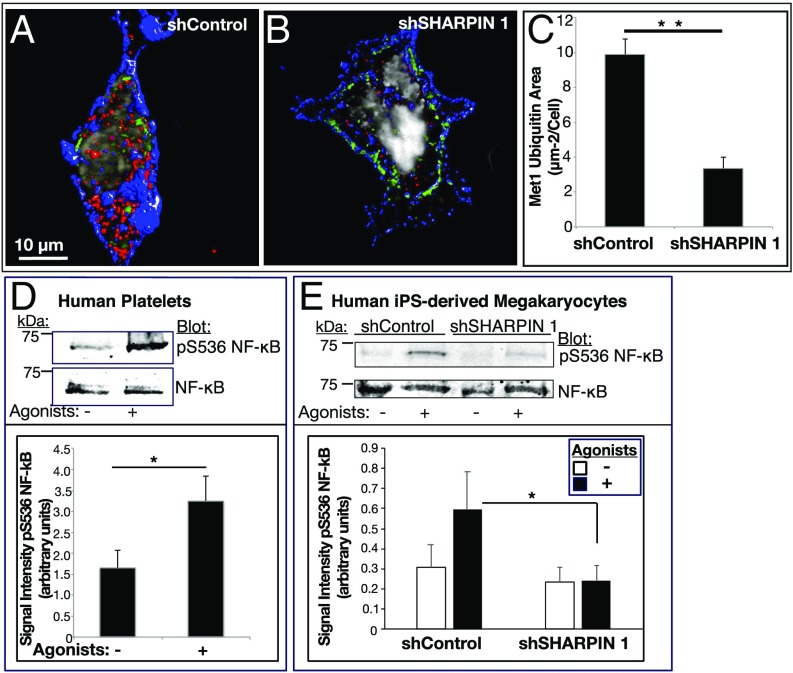
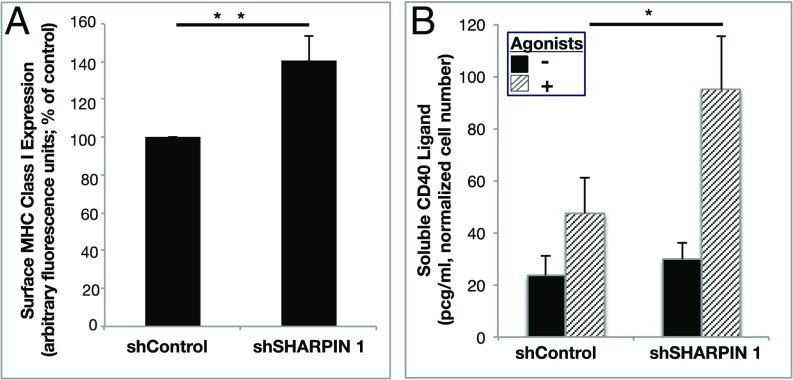
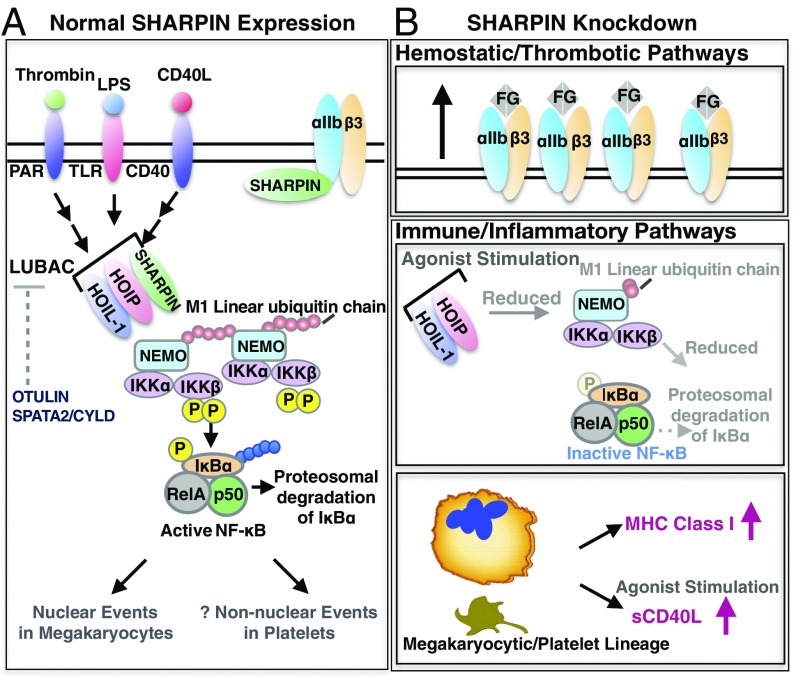
Platelets mediate primary hemostasis, and recent work has emphasized platelet participation in immunity and inflammation. The function of the platelet-specific integrin αIIbβ3 as a fibrinogen receptor in hemostasis is well defined, but the roles of αIIbβ3 or integrin-associated proteins in nonhemostatic platelet functions are poorly understood. Here we show that human platelets express the integrin-associated protein SHARPIN with functional consequences. In leukocytes, SHARPIN interacts with integrin α cytoplasmic tails, and it is also an obligate member of the linear ubiquitin chain assembly complex (LUBAC), which mediates Met1 linear ubiquitination of proteins leading to canonical NF-κB activation. SHARPIN interacted with αIIb in pull-down and coimmunoprecipitation assays. SHARPIN was partially localized, as was αIIbβ3, at platelet edges, and thrombin stimulation induced more central SHARPIN localization. SHARPIN also coimmunoprecipitated from platelets with the two other proteins comprising LUBAC, the E3 ligase HOIP and HOIL-1. Platelet stimulation with thrombin or inflammatory agonists, including lipopolysaccharide or soluble CD40 ligand (sCD40L), induced Met1 linear ubiquitination of the NF-κB pathway protein NEMO and serine-536 phosphorylation of the p65 RelA subunit of NF-κB. In human megakaryocytes and/or platelets derived from induced pluripotent stem (iPS) cells, SHARPIN knockdown caused increased basal and agonist-induced fibrinogen binding to αIIbβ3 as well as reduced Met1 ubiquitination and RelA phosphorylation. Moreover, these SHARPIN knockdown cells exhibited increased surface expression of MHC class I molecules and increased release of sCD40L. These results establish that SHARPIN functions in the human megakaryocyte/platelet lineage through protein interactions at the nexus of integrin and immune/inflammatory signaling.
Keywords: LUBAC; hemostasis; inflammation; integrin; platelet.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Platelet SHARPIN regulates platelet adhesion and inflammatory responses through associations with αIIbβ3 and LUBAC.Blood Adv. 2022 Apr 26;6(8):2595-2607. doi: 10.1182/bloodadvances.2021005611. Blood Adv. 2022. PMID: 34991155 Free PMC article.
-
Porcine Reproductive and Respiratory Syndrome Virus nsp1α Inhibits NF-κB Activation by Targeting the Linear Ubiquitin Chain Assembly Complex.J Virol. 2017 Jan 18;91(3):e01911-16. doi: 10.1128/JVI.01911-16. Print 2017 Feb 1. J Virol. 2017. PMID: 27881655 Free PMC article.
-
Genetic Instruction of Megakaryocytes and Platelets Derived from Human Induced Pluripotent Stem Cells for Studies of Integrin Regulation.Methods Mol Biol. 2021;2217:237-249. doi: 10.1007/978-1-0716-0962-0_13. Methods Mol Biol. 2021. PMID: 33215384
-
Linear ubiquitination: a novel NF-κB regulatory mechanism for inflammatory and immune responses by the LUBAC ubiquitin ligase complex.Endocr J. 2012;59(8):641-52. doi: 10.1507/endocrj.ej12-0148. Epub 2012 May 19. Endocr J. 2012. PMID: 22673407 Review.
-
Linear Ubiquitin Code: Its Writer, Erasers, Decoders, Inhibitors, and Implications in Disorders.Int J Mol Sci. 2020 May 11;21(9):3381. doi: 10.3390/ijms21093381. Int J Mol Sci. 2020. PMID: 32403254 Free PMC article. Review.
Cited by
-
Sharpin suppresses β1-integrin activation by complexing with the β1 tail and kindlin-1.Cell Commun Signal. 2019 Aug 20;17(1):101. doi: 10.1186/s12964-019-0407-6. Cell Commun Signal. 2019. PMID: 31429758 Free PMC article.
-
Cysteine-rich zinc finger proteins and the nuclear factor kappa-B pathway.Front Chem Biol. 2024;3:1503390. doi: 10.3389/fchbi.2024.1503390. Epub 2024 Dec 19. Front Chem Biol. 2024. PMID: 40405983 Free PMC article.
-
Targeting integrin pathways: mechanisms and advances in therapy.Signal Transduct Target Ther. 2023 Jan 2;8(1):1. doi: 10.1038/s41392-022-01259-6. Signal Transduct Target Ther. 2023. PMID: 36588107 Free PMC article. Review.
-
Integrin Binding Dynamics Modulate Ligand-Specific Mechanosensing in Mammary Gland Fibroblasts.iScience. 2020 Mar 27;23(3):100907. doi: 10.1016/j.isci.2020.100907. Epub 2020 Feb 13. iScience. 2020. PMID: 32106057 Free PMC article.
-
Genome-Wide Association Studies for the Concentration of Albumin in Colostrum and Serum in Chinese Holstein.Animals (Basel). 2020 Nov 26;10(12):2211. doi: 10.3390/ani10122211. Animals (Basel). 2020. PMID: 33255903 Free PMC article.
References
-
- Bennett JS. Regulation of integrins in platelets. Biopolymers. 2015;104:323–333. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
