

Tissue-specific BMAL1 cistromes reveal that rhythmic transcription is associated with rhythmic enhancer-enhancer interactions
- PMID: 30804225
- PMCID: PMC6411008
- DOI: 10.1101/gad.322198.118
Tissue-specific BMAL1 cistromes reveal that rhythmic transcription is associated with rhythmic enhancer-enhancer interactions
Abstract
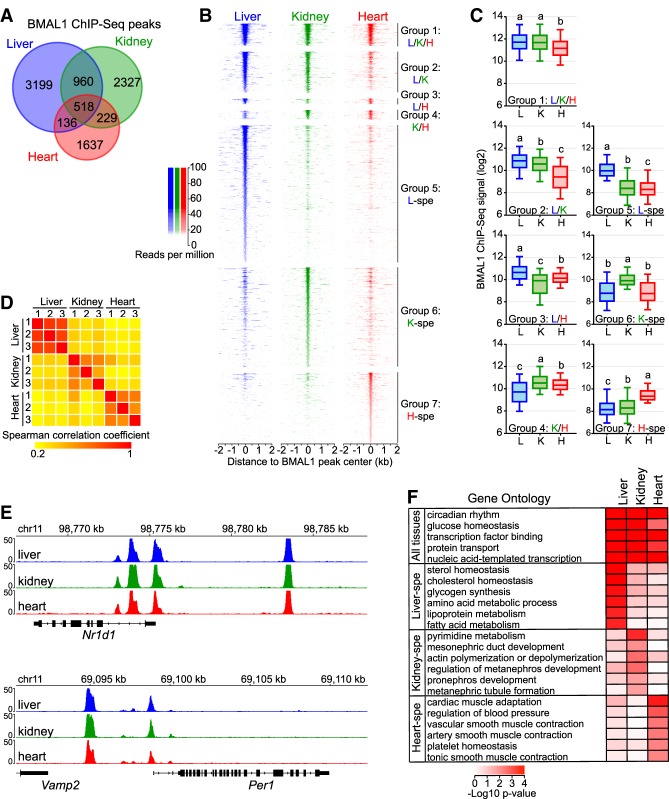
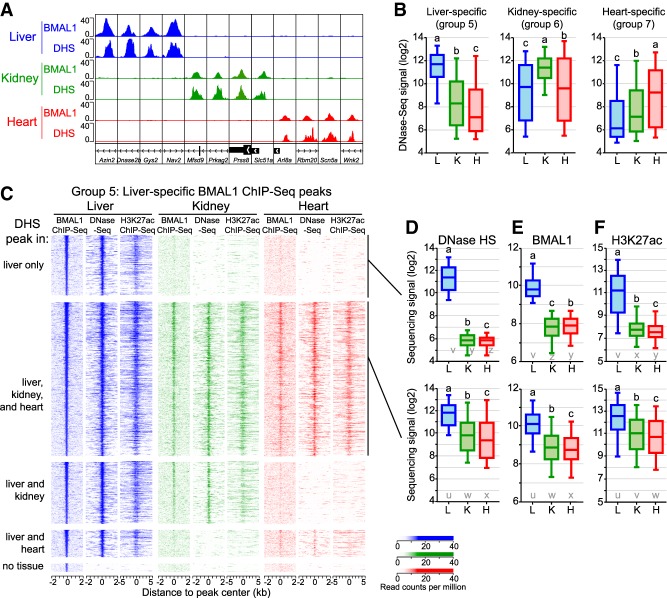
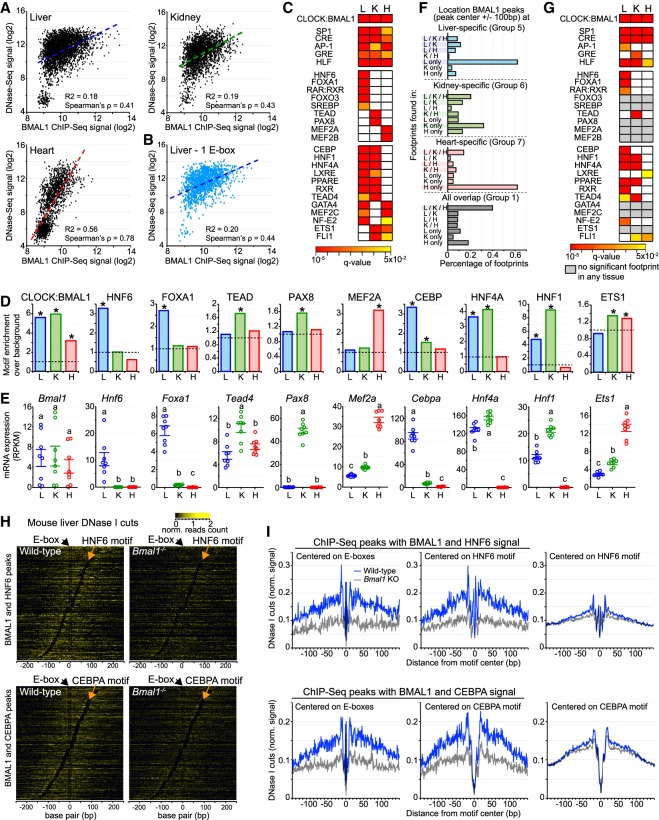
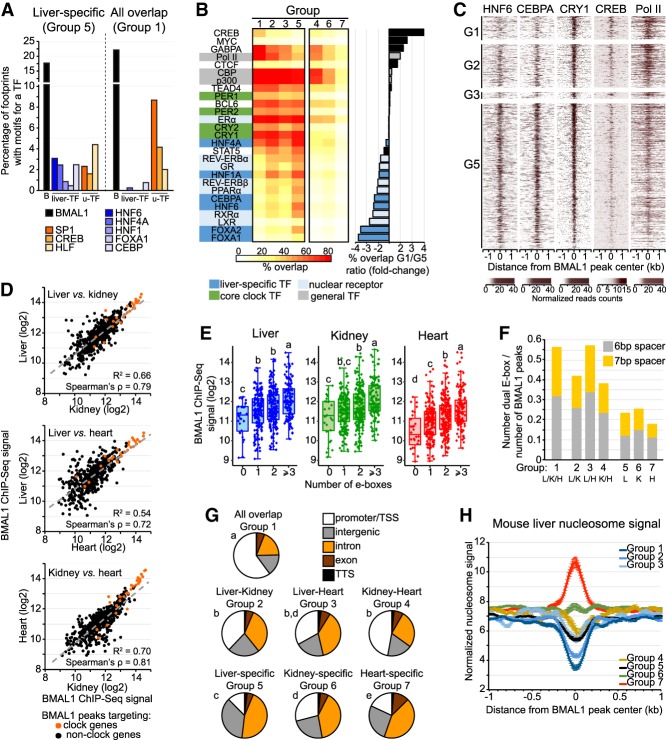
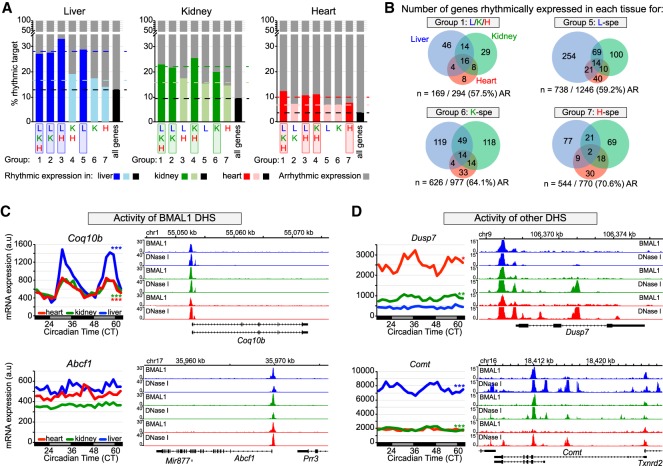
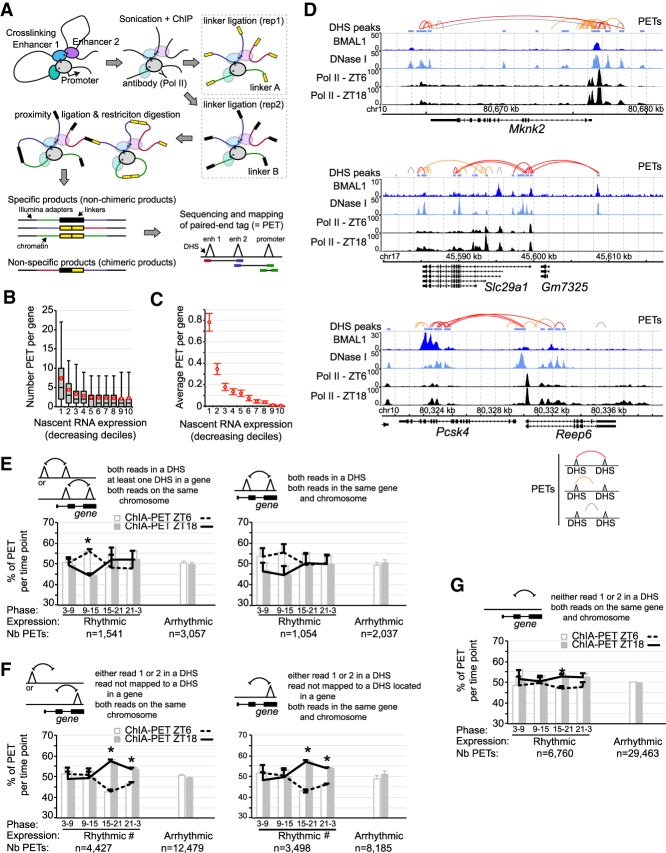
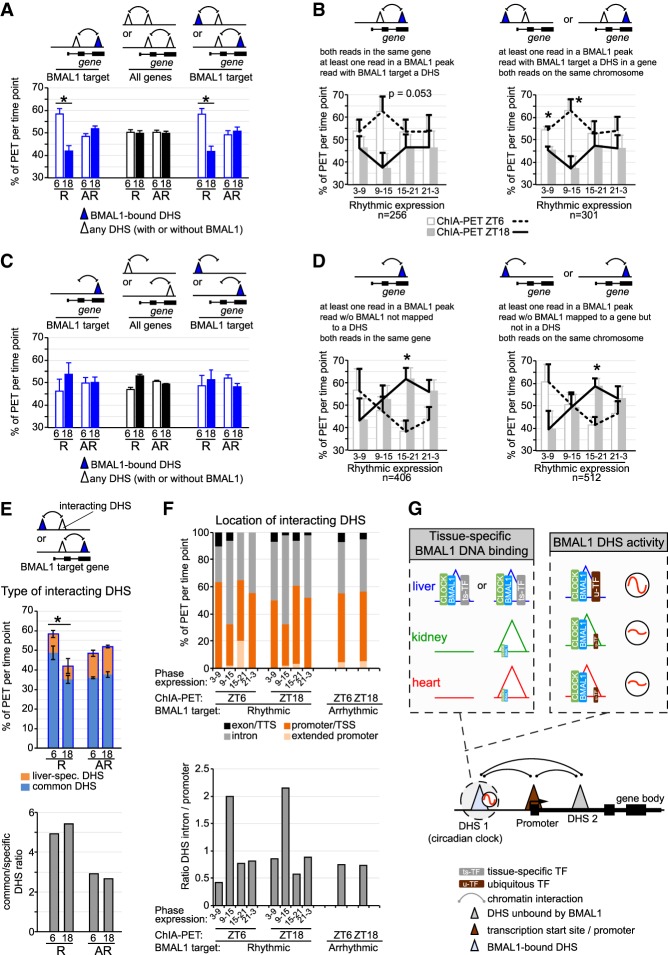
The mammalian circadian clock relies on the transcription factor CLOCK:BMAL1 to coordinate the rhythmic expression of thousands of genes. Consistent with the various biological functions under clock control, rhythmic gene expression is tissue-specific despite an identical clockwork mechanism in every cell. Here we show that BMAL1 DNA binding is largely tissue-specific, likely because of differences in chromatin accessibility between tissues and cobinding of tissue-specific transcription factors. Our results also indicate that BMAL1 ability to drive tissue-specific rhythmic transcription is associated with not only the activity of BMAL1-bound enhancers but also the activity of neighboring enhancers. Characterization of physical interactions between BMAL1 enhancers and other cis-regulatory regions by RNA polymerase II chromatin interaction analysis by paired-end tag (ChIA-PET) reveals that rhythmic BMAL1 target gene expression correlates with rhythmic chromatin interactions. These data thus support that much of BMAL1 target gene transcription depends on BMAL1 capacity to rhythmically regulate a network of enhancers.
Keywords: circadian clock; enhancer–enhancer interactions; tissue-specific cistromes; transcription.
© 2019 Beytebiere et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
Comment in
-
Help from my friends-cooperation of BMAL1 with noncircadian transcription factors.Genes Dev. 2019 Mar 1;33(5-6):255-257. doi: 10.1101/gad.324046.119. Genes Dev. 2019. PMID: 30824531 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases