

The PD-1/PD-L1 Pathway Affects the Expansion and Function of Cytotoxic CD8+ T Cells During an Acute Retroviral Infection
- PMID: 30804928
- PMCID: PMC6370637
- DOI: 10.3389/fimmu.2019.00054
The PD-1/PD-L1 Pathway Affects the Expansion and Function of Cytotoxic CD8+ T Cells During an Acute Retroviral Infection
Abstract
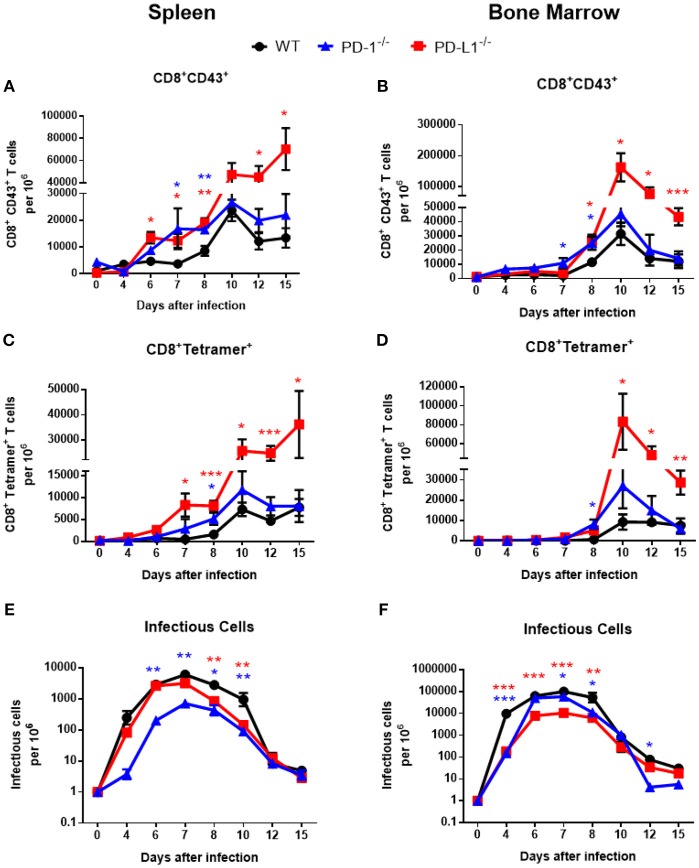
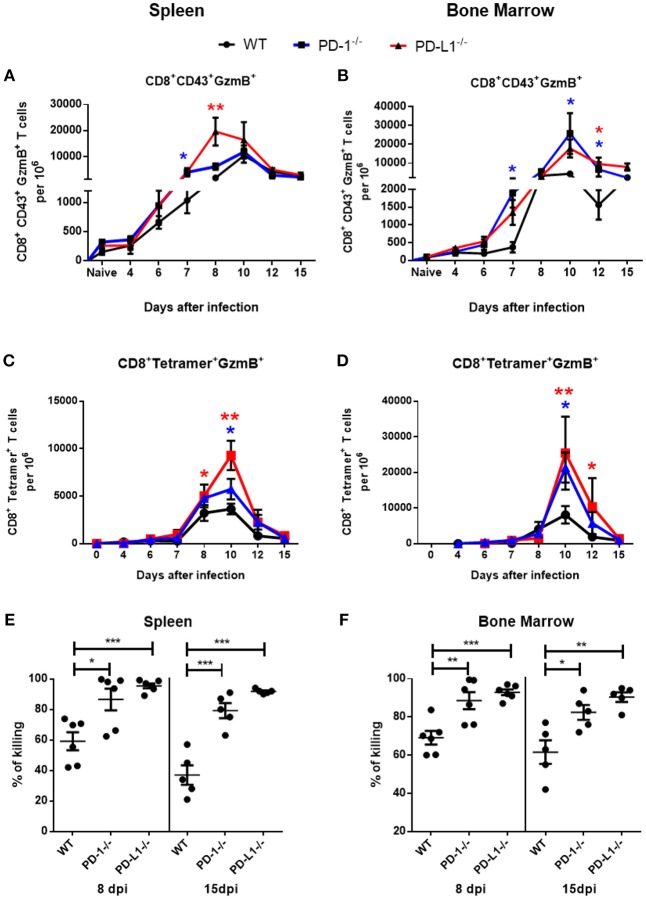
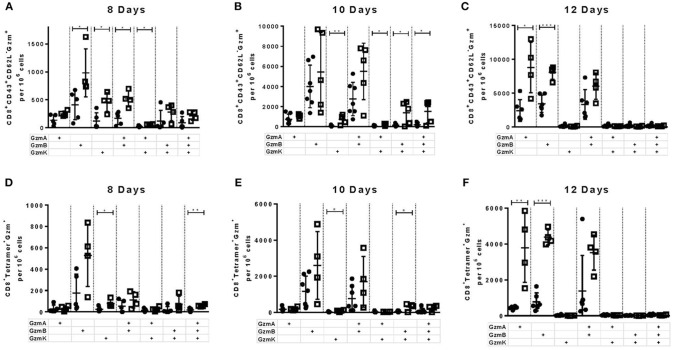
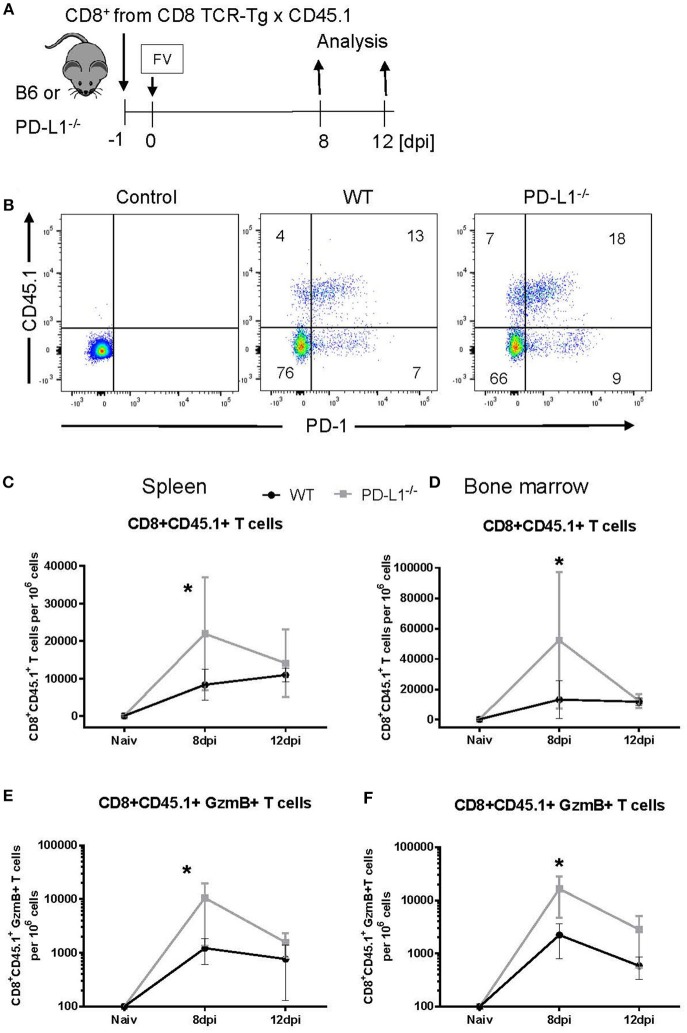
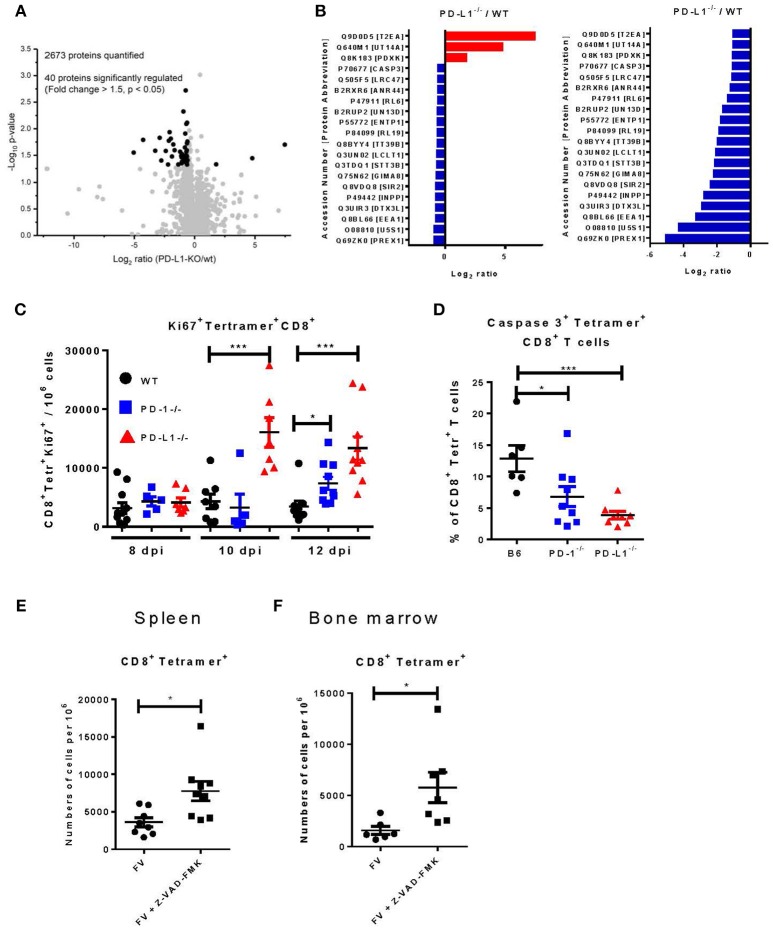
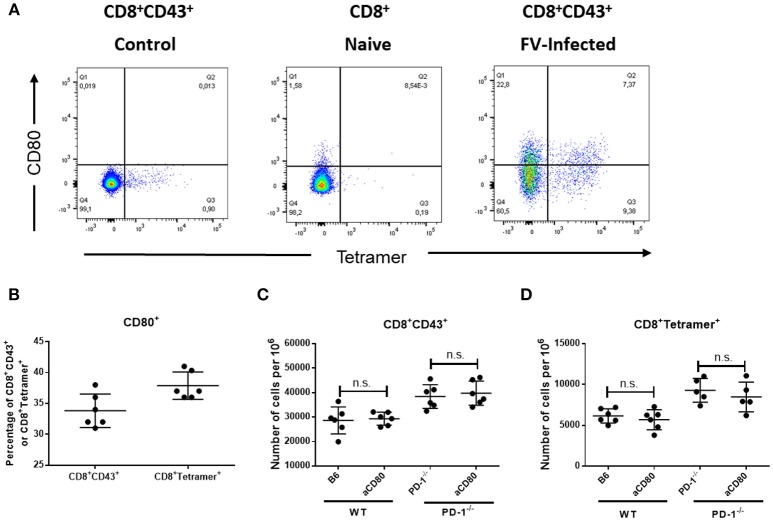
Cytotoxic CD8+ T lymphocytes (CTL) efficiently control acute virus infections but can become exhausted when a chronic infection develops. The checkpoint receptor PD-1 suppresses the functionality of virus-specific CD8+ T cells during chronic infection. However, the role of the PD-L1/PD-1 pathway during the acute phase of infections has not been well characterized. In the current study the effects of PD-1 or PD-L1 deficiency on the CD8+ T cell response against Friend retroviral (FV) infection of knockout mice was analyzed during acute infection. We observed an enhanced proliferation, functional maturation, and reduced apoptosis of effector CD8+ T cells in the absence of PD-1 or PD-L1. The knockout of PD-L1 had a stronger effect on the functionality of CD8+ T cells than that of PD-1. Augmented CTL responses were associated with an improved control of FV replication. The strong phenotype of FV-infected PD-L1 knockout mice was independent of the interaction with CD80 as an additional receptor for PD-L1. Furthermore, we performed a detailed analysis of the production of different granzymes in virus-specific CD8+ T cells and observed that especially the simultaneous production of multiple granzymes in individual T cells (multifunctionality) was under the control of the PD-1/PD-L1 pathway. The findings from this study allow for a better understanding of the development of antiviral cytotoxic immunity during acute viral infections.
Keywords: CD8 T cells; PD-1; PD-L1; apoptosis; caspase 3; immunoregulation; retrovirus.
Figures
Similar articles
-
PD-L1 Expression on Retrovirus-Infected Cells Mediates Immune Escape from CD8+ T Cell Killing.PLoS Pathog. 2015 Oct 20;11(10):e1005224. doi: 10.1371/journal.ppat.1005224. eCollection 2015 Oct. PLoS Pathog. 2015. PMID: 26484769 Free PMC article.
-
PD-1 knockout on cytotoxic primary murine CD8+ T cells improves their motility in retrovirus infected mice.Front Immunol. 2024 Apr 29;15:1338218. doi: 10.3389/fimmu.2024.1338218. eCollection 2024. Front Immunol. 2024. PMID: 38742109 Free PMC article.
-
A Combination of Anti-PD-L1 Treatment and Therapeutic Vaccination Facilitates Improved Retroviral Clearance via Reactivation of Highly Exhausted T Cells.mBio. 2021 Feb 2;12(1):e02121-20. doi: 10.1128/mBio.02121-20. mBio. 2021. PMID: 33531395 Free PMC article.
-
The PD-1/PD-L1 (B7-H1) pathway in chronic infection-induced cytotoxic T lymphocyte exhaustion.J Biomed Biotechnol. 2011;2011:451694. doi: 10.1155/2011/451694. Epub 2011 Sep 25. J Biomed Biotechnol. 2011. PMID: 21960736 Free PMC article. Review.
-
Costimulatory molecule programmed death-1 in the cytotoxic response during chronic hepatitis C.World J Gastroenterol. 2009 Nov 7;15(41):5129-40. doi: 10.3748/wjg.15.5129. World J Gastroenterol. 2009. PMID: 19891011 Free PMC article. Review.
Cited by
-
Combination rhIL-15 and Anti-PD-L1 (Avelumab) Enhances HIVGag-Specific CD8 T-Cell Function.J Infect Dis. 2020 Oct 1;222(9):1540-1549. doi: 10.1093/infdis/jiaa269. J Infect Dis. 2020. PMID: 32433762 Free PMC article.
-
Regulatory T cells suppress the motility of cytotoxic T cells in Friend retrovirus-infected mice.JCI Insight. 2023 Jul 10;8(13):e167482. doi: 10.1172/jci.insight.167482. JCI Insight. 2023. PMID: 37427590 Free PMC article.
-
Crosstalk between Apoptosis and Cytotoxic Lymphocytes (CTLs) in the Course of Lagovirus Europaeus GI.1a Infection in Rabbits.J Vet Res. 2023 Feb 28;67(1):41-47. doi: 10.2478/jvetres-2023-0008. eCollection 2023 Mar. J Vet Res. 2023. PMID: 37008759 Free PMC article.
-
Bioinformatic analysis of PD-1 checkpoint blockade response in influenza infection.BMC Genom Data. 2022 Aug 13;23(1):65. doi: 10.1186/s12863-022-01081-7. BMC Genom Data. 2022. PMID: 35962325 Free PMC article.
-
Distribution characteristics of serum β2-microglobulin between viral and bacterial lower respiratory tract infections: a retrospective study.PeerJ. 2020 Aug 25;8:e9814. doi: 10.7717/peerj.9814. eCollection 2020. PeerJ. 2020. PMID: 32904447 Free PMC article.
References
-
- Zelinskyy G, Myers L, Dietze KK, Gibbert K, Roggendorf M, Liu J, et al. . Virus-specific CD8+ T cells upregulate programmed death-1 expression during acute friend retrovirus infection but are highly cytotoxic and control virus replication. J Immunol. (2011) 187:3730–7. 10.4049/jimmunol.1101612 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials