

A Model of Motion Processing in the Visual Cortex Using Neural Field With Asymmetric Hebbian Learning
- PMID: 30809113
- PMCID: PMC6380226
- DOI: 10.3389/fnins.2019.00067
A Model of Motion Processing in the Visual Cortex Using Neural Field With Asymmetric Hebbian Learning
Abstract
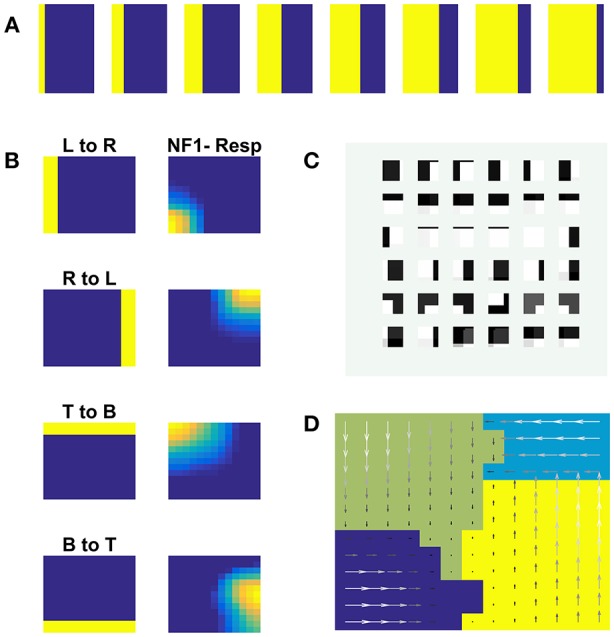
Neurons in the dorsal pathway of the visual cortex are thought to be involved in motion processing. The first site of motion processing is the primary visual cortex (V1), encoding the direction of motion in local receptive fields, with higher order motion processing happening in the middle temporal area (MT). Complex motion properties like optic flow are processed in higher cortical areas of the Medial Superior Temporal area (MST). In this study, a hierarchical neural field network model of motion processing is presented. The model architecture has an input layer followed by either one or cascade of two neural fields (NF): the first of these, NF1, represents V1, while the second, NF2, represents MT. A special feature of the model is that lateral connections used in the neural fields are trained by asymmetric Hebbian learning, imparting to the neural field the ability to process sequential information in motion stimuli. The model was trained using various traditional moving patterns such as bars, squares, gratings, plaids, and random dot stimulus. In the case of bar stimuli, the model had only a single NF, the neurons of which developed a direction map of the moving bar stimuli. Training a network with two NFs on moving square and moving plaids stimuli, we show that, while the neurons in NF1 respond to the direction of the component (such as gratings and edges) motion, the neurons in NF2 (analogous to MT) responding to the direction of the pattern (plaids, square object) motion. In the third study, a network with 2 NFs was simulated using random dot stimuli (RDS) with translational motion, and show that the NF2 neurons can encode the direction of the concurrent dot motion (also called translational flow motion), independent of the dot configuration. This translational RDS flow motion is decoded by a simple perceptron network (a layer above NF2) with an accuracy of 100% on train set and 90% on the test set, thereby demonstrating that the proposed network can generalize to new dot configurations. Also, the response properties of the model on different input stimuli closely resembled many of the known features of the neurons found in electrophysiological studies.
Keywords: lateral interactions; medial superior temporal area (MST); middle temporal area (MT); neural field models; pattern selectivity; primary visual area (V1); weight asymmetry.
Figures









Similar articles
-
Responses of MT and MST neurons to one and two moving objects in the receptive field.J Neurophysiol. 1997 Dec;78(6):2904-15. doi: 10.1152/jn.1997.78.6.2904. J Neurophysiol. 1997. PMID: 9405511
-
Direction and orientation selectivity of neurons in visual area MT of the macaque.J Neurophysiol. 1984 Dec;52(6):1106-30. doi: 10.1152/jn.1984.52.6.1106. J Neurophysiol. 1984. PMID: 6520628
-
Temporal and spatial limits of pattern motion sensitivity in macaque MT neurons.J Neurophysiol. 2015 Apr 1;113(7):1977-88. doi: 10.1152/jn.00597.2014. Epub 2014 Dec 24. J Neurophysiol. 2015. PMID: 25540222 Free PMC article.
-
A neural-based code for computing image velocity from small sets of middle temporal (MT/V5) neuron inputs.J Vis. 2012 Aug 1;12(8):1. doi: 10.1167/12.8.1. J Vis. 2012. PMID: 22854102 Review.
-
Catching the voltage gradient-asymmetric boost of cortical spread generates motion signals across visual cortex: a brief review with special thanks to Amiram Grinvald.Neurophotonics. 2017 Jul;4(3):031206. doi: 10.1117/1.NPh.4.3.031206. Epub 2017 Feb 10. Neurophotonics. 2017. PMID: 28217713 Free PMC article. Review.
Cited by
-
Modeling the development of cortical responses in primate dorsal ("where") pathway to optic flow using hierarchical neural field models.Front Neurosci. 2023 May 22;17:1154252. doi: 10.3389/fnins.2023.1154252. eCollection 2023. Front Neurosci. 2023. PMID: 37284658 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous