

Expression of the Phosphatase Ppef2 Controls Survival and Function of CD8+ Dendritic Cells
- PMID: 30809231
- PMCID: PMC6379467
- DOI: 10.3389/fimmu.2019.00222
Expression of the Phosphatase Ppef2 Controls Survival and Function of CD8+ Dendritic Cells
Abstract
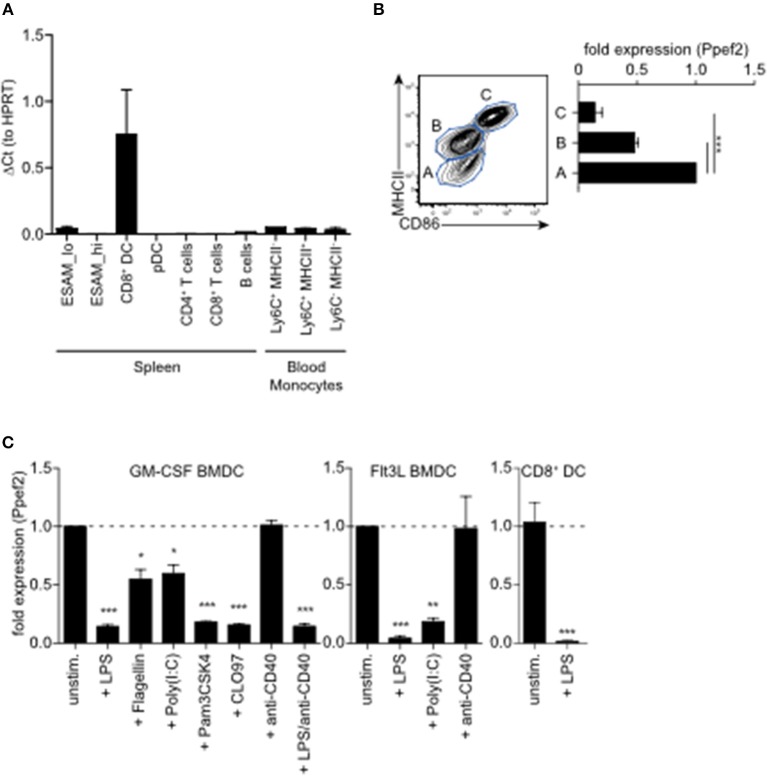
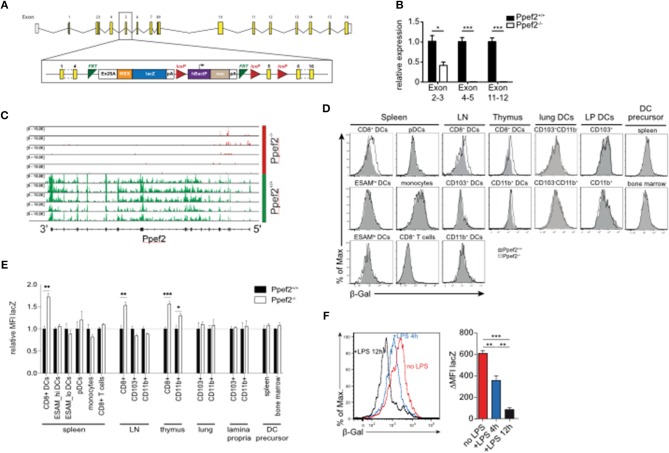
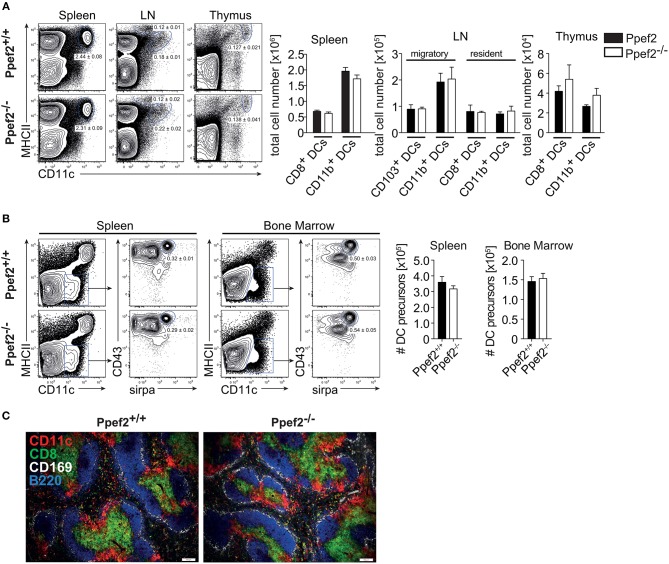
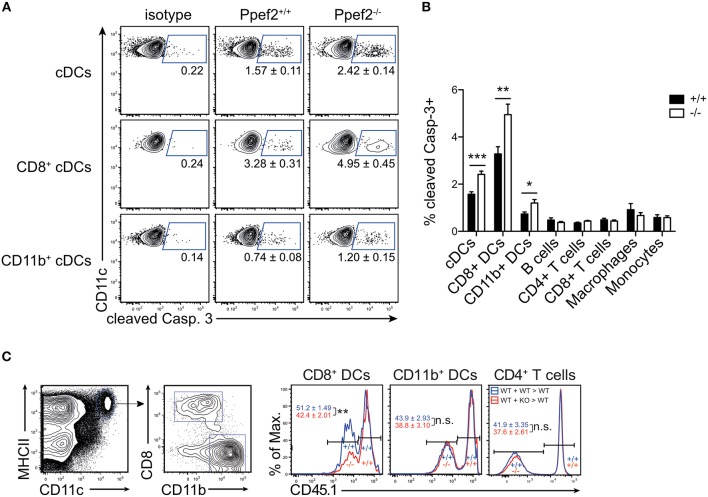
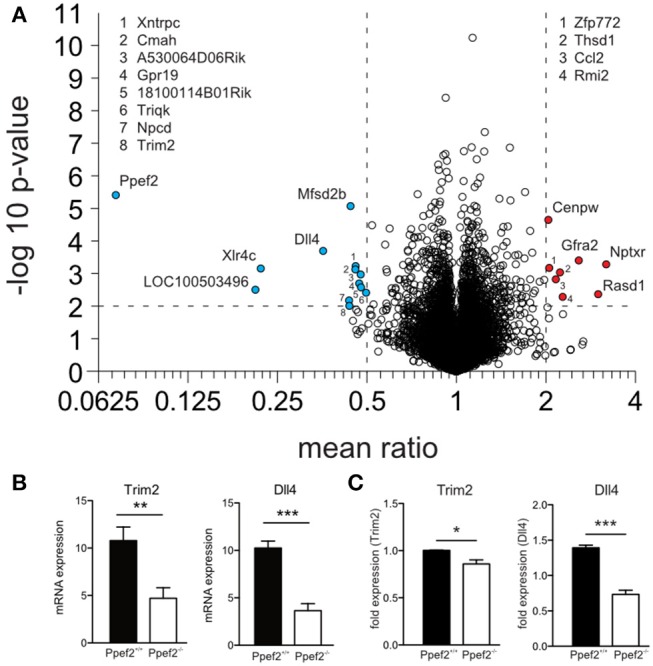
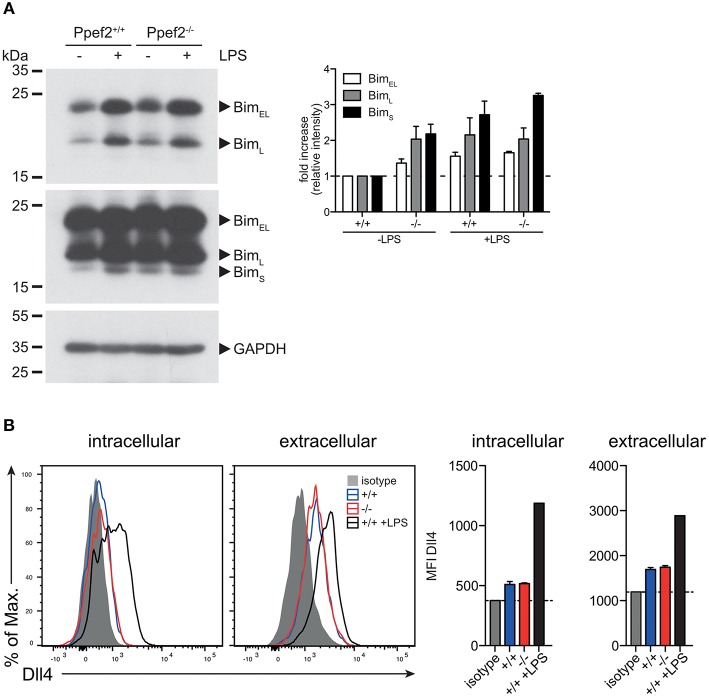
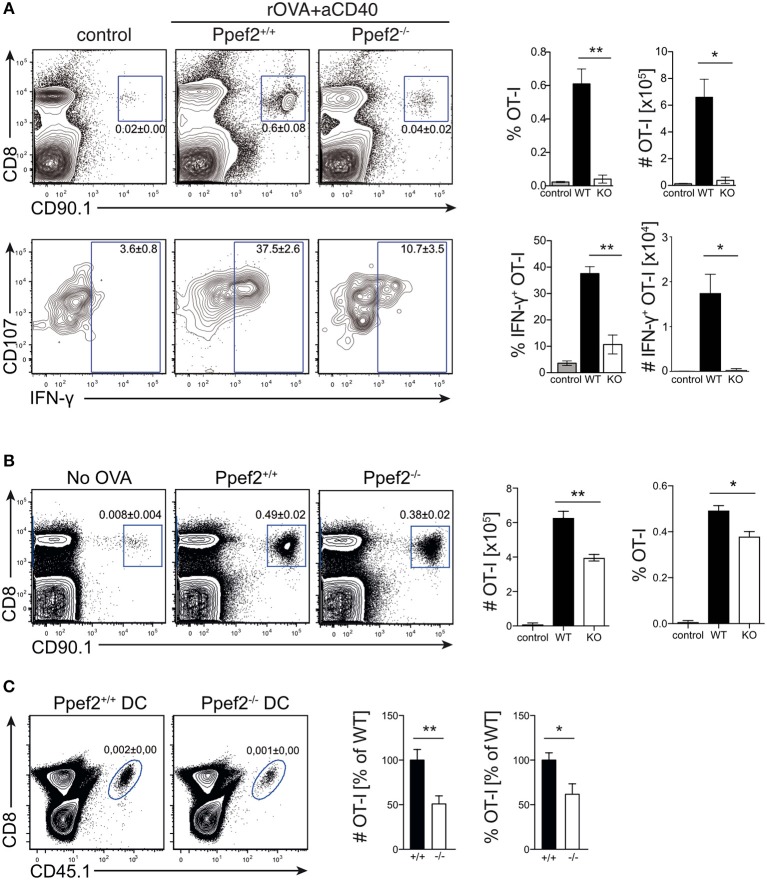
Apoptotic cell death of Dendritic cells (DCs) is critical for immune homeostasis. Although intrinsic mechanisms controlling DC death have not been fully characterized up to now, experimentally enforced inhibition of DC-death causes various autoimmune diseases in model systems. We have generated mice deficient for Protein Phosphatase with EF-Hands 2 (Ppef2), which is selectively expressed in CD8+ DCs, but not in other related DC subtypes such as tissue CD103+ DCs. Ppef2 is down-regulated rapidly upon maturation of DCs by toll-like receptor stimuli, but not upon triggering of CD40. Ppef2-deficient CD8+ DCs accumulate the pro-apoptotic Bcl-2-like protein 11 (Bim) and show increased apoptosis and reduced competitve repopulation capacities. Furthermore, Ppef2-/- CD8+ DCs have strongly diminished antigen presentation capacities in vivo, as CD8+ T cells primed by Ppef2-/- CD8+ DCs undergo reduced expansion. In conclusion, our data suggests that Ppef2 is crucial to support survival of immature CD8+ DCs, while Ppef2 down-regulation during DC-maturation limits T cell responses.
Keywords: CD8 T cell priming; DC-maturation; apoptosis; cross-presentation; dendritic cells; immune homeostasis.
Figures
Similar articles
-
Outer membrane vesicles engineered to express membrane-bound antigen program dendritic cells for cross-presentation to CD8+ T cells.Acta Biomater. 2019 Jun;91:248-257. doi: 10.1016/j.actbio.2019.04.033. Epub 2019 Apr 17. Acta Biomater. 2019. PMID: 31003032
-
Enhancement of dendritic cell-based vaccine potency by anti-apoptotic siRNAs targeting key pro-apoptotic proteins in cytotoxic CD8(+) T cell-mediated cell death.Immunol Lett. 2009 Jan 29;122(1):58-67. doi: 10.1016/j.imlet.2008.12.006. Epub 2009 Jan 9. Immunol Lett. 2009. PMID: 19135479
-
Prostaglandin E2 in a TLR3- and 7/8-agonist-based DC maturation cocktail generates mature, cytokine-producing, migratory DCs but impairs antigen cross-presentation to CD8+ T cells.Cancer Immunol Immunother. 2020 Jun;69(6):1029-1042. doi: 10.1007/s00262-019-02470-1. Epub 2020 Feb 25. Cancer Immunol Immunother. 2020. PMID: 32100075 Free PMC article.
-
Glycan-based DC-SIGN targeting vaccines to enhance antigen cross-presentation.Mol Immunol. 2013 Sep;55(2):143-5. doi: 10.1016/j.molimm.2012.10.031. Epub 2012 Nov 14. Mol Immunol. 2013. PMID: 23158834 Review.
-
Unique functions of splenic CD8alpha+ dendritic cells during infection with intracellular pathogens.Immunol Lett. 2007 Dec 15;114(2):66-72. doi: 10.1016/j.imlet.2007.09.007. Epub 2007 Oct 12. Immunol Lett. 2007. PMID: 17964665 Review.
Cited by
-
GM-CSF, Flt3-L and IL-4 affect viability and function of conventional dendritic cell types 1 and 2.Front Immunol. 2023 Jan 12;13:1058963. doi: 10.3389/fimmu.2022.1058963. eCollection 2022. Front Immunol. 2023. PMID: 36713392 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials