

Aerobic Glycolysis Is Required for Spatial Memory Acquisition But Not Memory Retrieval in Mice
- PMID: 30809587
- PMCID: PMC6390195
- DOI: 10.1523/ENEURO.0389-18.2019
Aerobic Glycolysis Is Required for Spatial Memory Acquisition But Not Memory Retrieval in Mice
Abstract
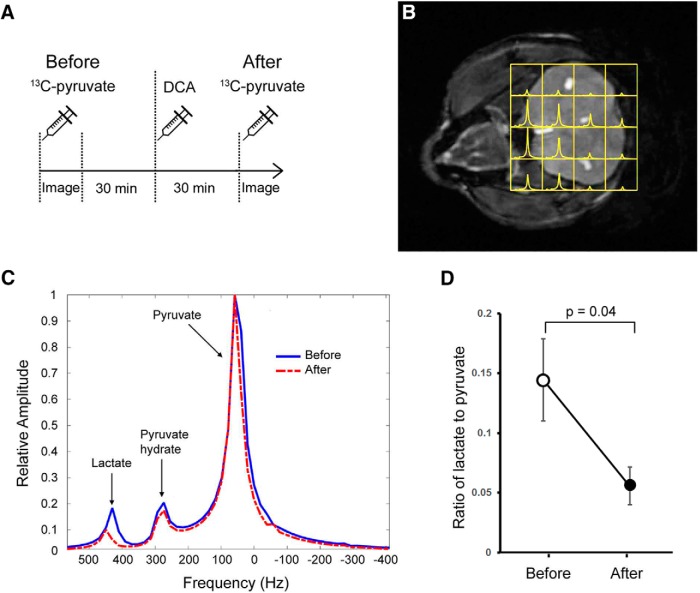
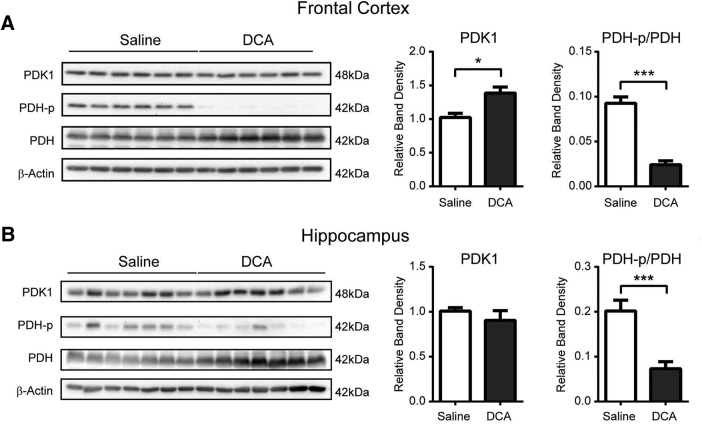
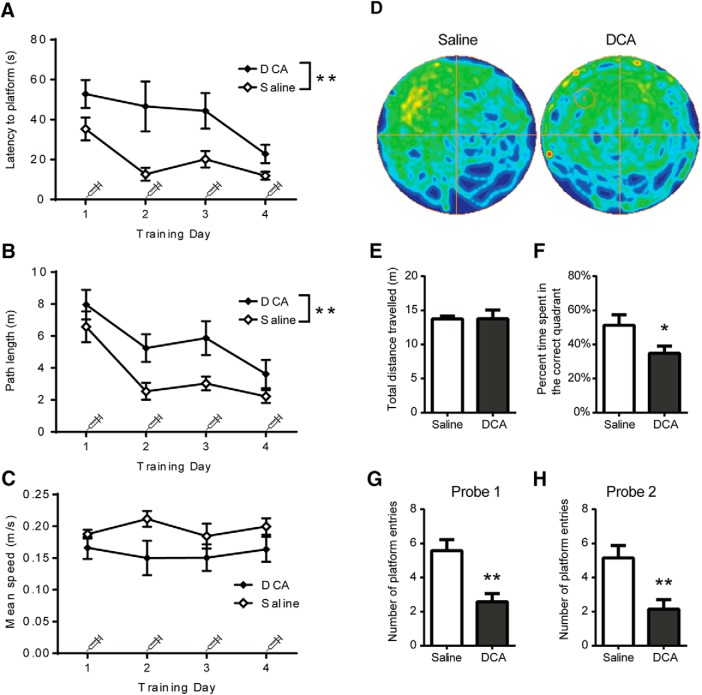
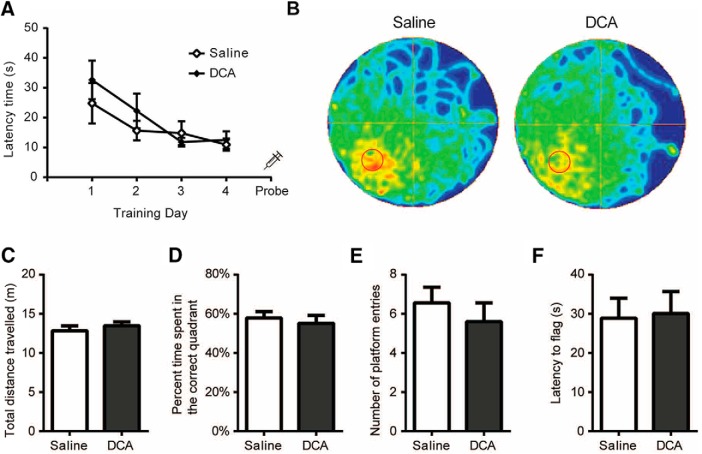
The consolidation of newly formed memories and their retrieval are energetically demanding processes. Aerobic glycolysis (AG), also known as the Warburg effect, consists of the production of lactate from glucose in the presence of oxygen. The astrocyte neuron lactate shuttle hypothesis posits that astrocytes process glucose by AG to generate lactate, which is used as a fuel source within neurons to maintain synaptic activity. Studies in mice have demonstrated that lactate transport between astrocytes and neurons is required for long-term memory formation, yet the role of lactate production in memory acquisition and retrieval has not previously been explored. Here, we examined the effect of dichloroacetate (DCA), a chemical inhibitor of lactate production, on spatial learning and memory in mice using the Morris water maze (MWM). In vivo hyperpolarized 13C-pyruvate magnetic resonance spectroscopy revealed decreased conversion of pyruvate to lactate in the mouse brain following DCA administration, concomitant with a reduction in the phosphorylation of pyruvate dehydrogenase. DCA exposure before each training session in the MWM impaired learning, which subsequently resulted in impaired memory during the probe trial. In contrast, mice that underwent training without DCA exposure, but received a single DCA injection before the probe trial exhibited normal memory. Our findings indicate that AG plays a key role during memory acquisition but is less important for the retrieval of established memories. Thus, the activation of AG may be important for learning-dependent synaptic plasticity rather than the activation of signaling cascades required for memory retrieval.
Keywords: aerobic glycolysis; lactate; magnetic resonance spectroscopy; memory; metabolism; synaptic plasticity.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources