

The moonlighting RNA-binding activity of cytosolic serine hydroxymethyltransferase contributes to control compartmentalization of serine metabolism
- PMID: 30809670
- PMCID: PMC6486632
- DOI: 10.1093/nar/gkz129
The moonlighting RNA-binding activity of cytosolic serine hydroxymethyltransferase contributes to control compartmentalization of serine metabolism
Abstract
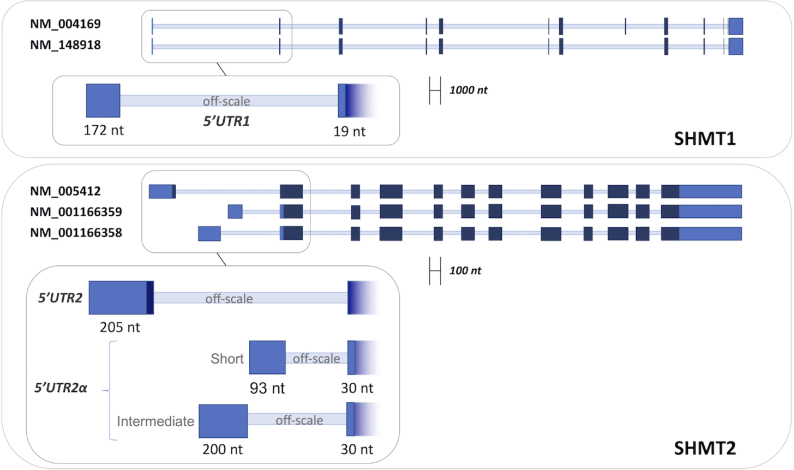
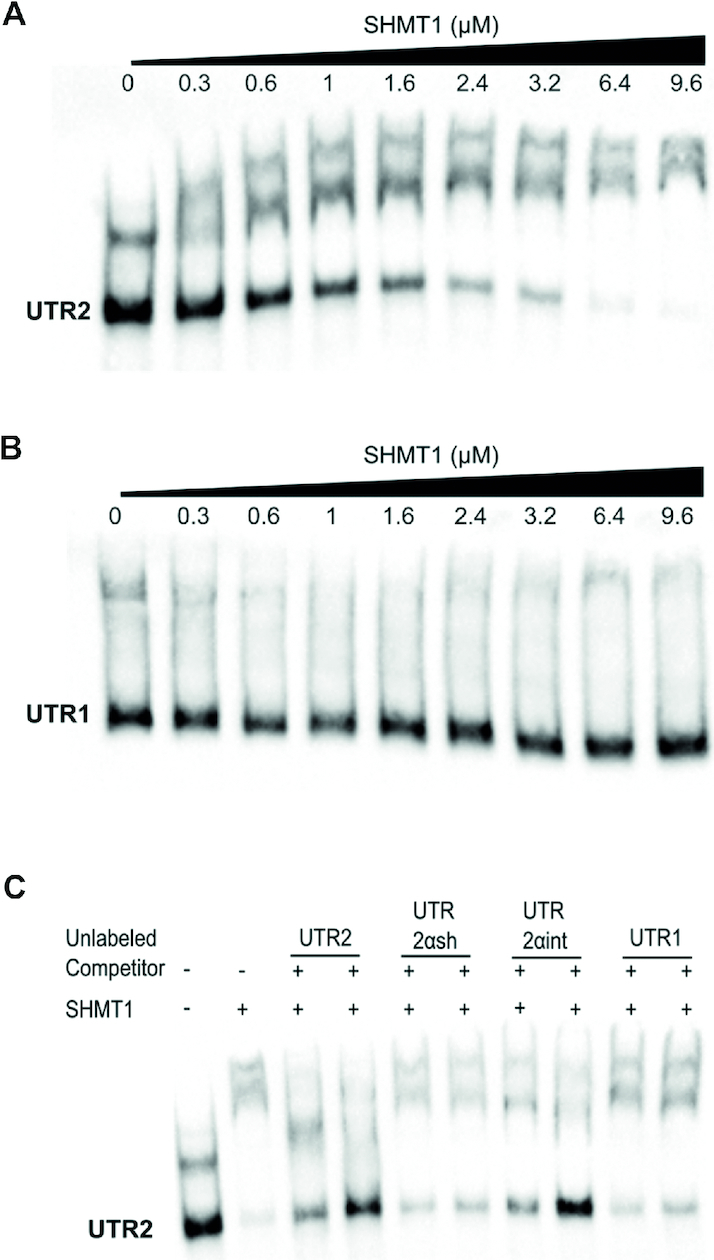
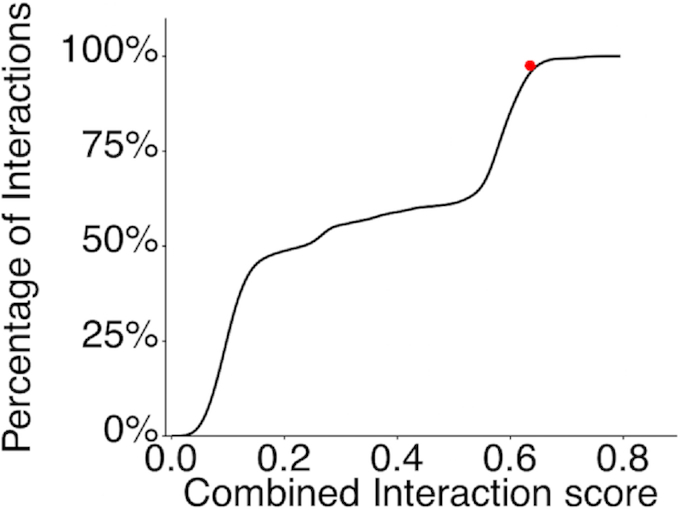
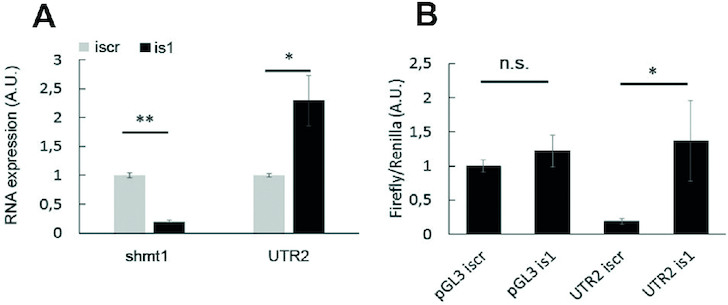
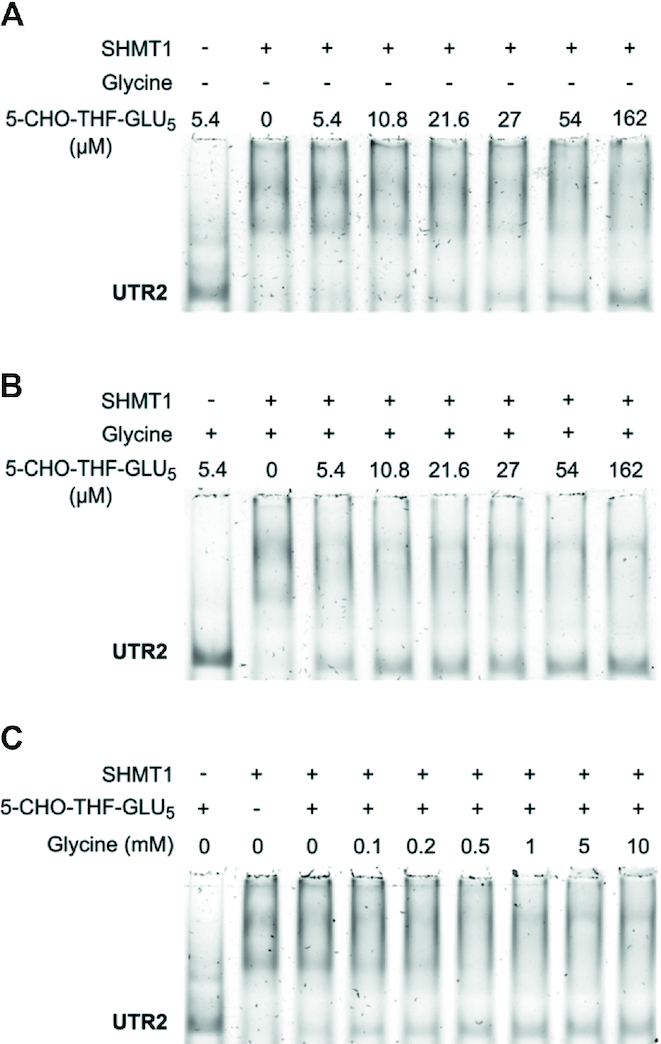
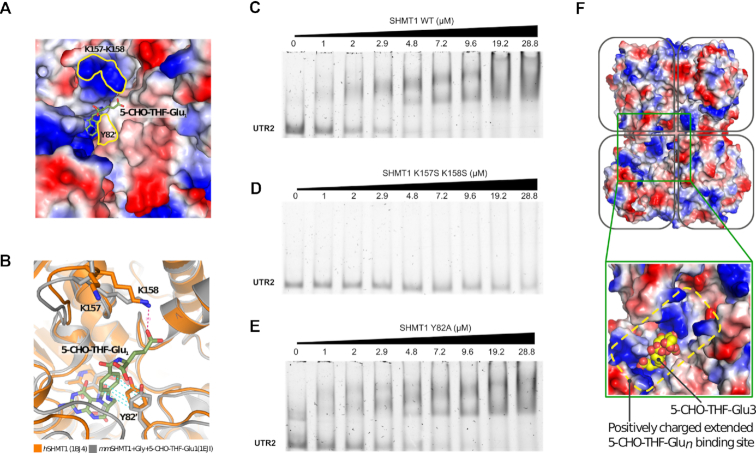
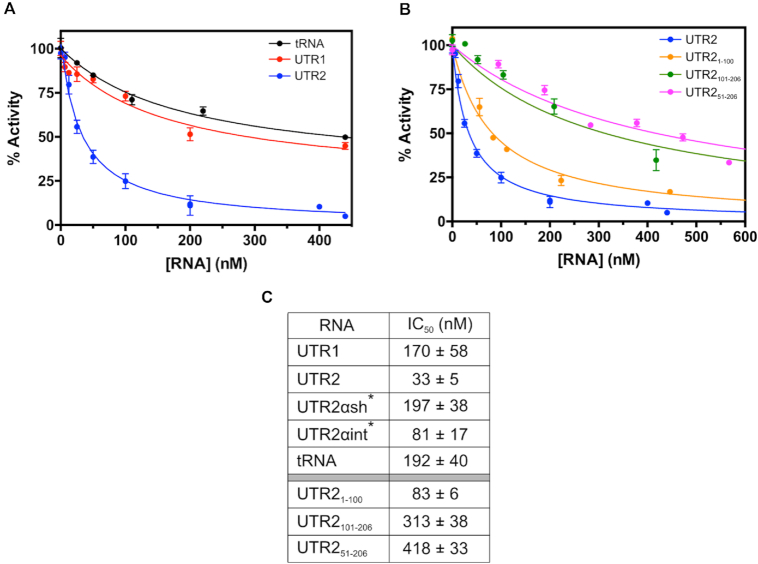
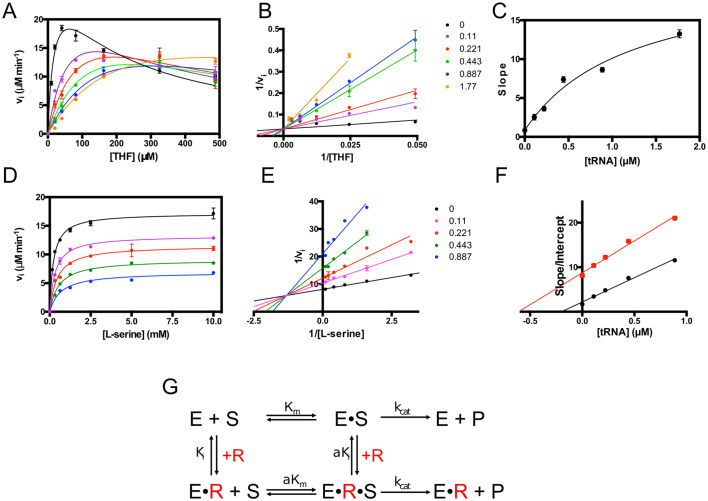
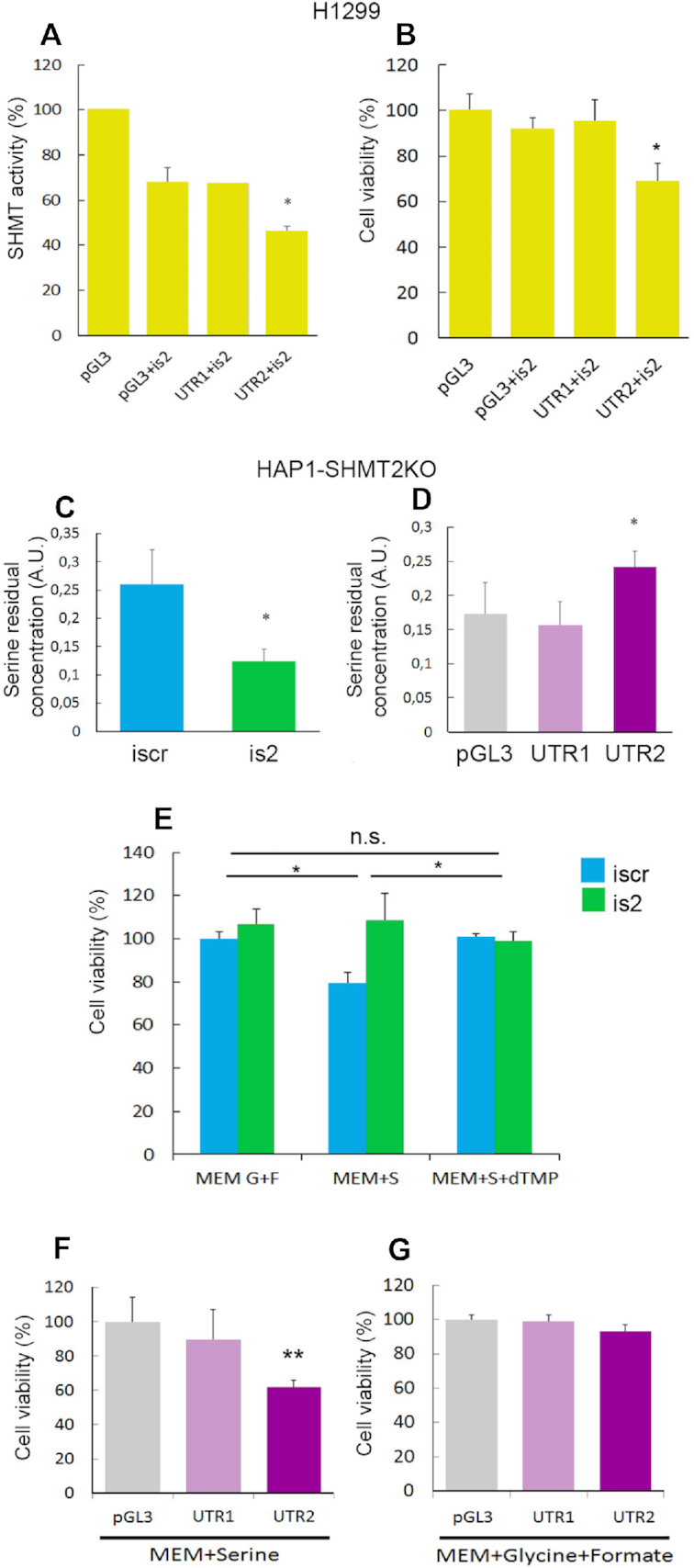
Enzymes of intermediary metabolism are often reported to have moonlighting functions as RNA-binding proteins and have regulatory roles beyond their primary activities. Human serine hydroxymethyltransferase (SHMT) is essential for the one-carbon metabolism, which sustains growth and proliferation in normal and tumour cells. Here, we characterize the RNA-binding function of cytosolic SHMT (SHMT1) in vitro and using cancer cell models. We show that SHMT1 controls the expression of its mitochondrial counterpart (SHMT2) by binding to the 5'untranslated region of the SHMT2 transcript (UTR2). Importantly, binding to RNA is modulated by metabolites in vitro and the formation of the SHMT1-UTR2 complex inhibits the serine cleavage activity of the SHMT1, without affecting the reverse reaction. Transfection of UTR2 in cancer cells controls SHMT1 activity and reduces cell viability. We propose a novel mechanism of SHMT regulation, which interconnects RNA and metabolites levels to control the cross-talk between cytosolic and mitochondrial compartments of serine metabolism.
© The Author(s) 2019. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
Differential 3-bromopyruvate inhibition of cytosolic and mitochondrial human serine hydroxymethyltransferase isoforms, key enzymes in cancer metabolic reprogramming.Biochim Biophys Acta. 2016 Nov;1864(11):1506-17. doi: 10.1016/j.bbapap.2016.08.010. Epub 2016 Aug 13. Biochim Biophys Acta. 2016. PMID: 27530298
-
Human Cytosolic and Mitochondrial Serine Hydroxymethyltransferase Isoforms in Comparison: Full Kinetic Characterization and Substrate Inhibition Properties.Biochemistry. 2018 Dec 26;57(51):6984-6996. doi: 10.1021/acs.biochem.8b01074. Epub 2018 Dec 7. Biochemistry. 2018. PMID: 30500180
-
The catalytic activity of serine hydroxymethyltransferase is essential for de novo nuclear dTMP synthesis in lung cancer cells.FEBS J. 2018 Sep;285(17):3238-3253. doi: 10.1111/febs.14610. Epub 2018 Aug 7. FEBS J. 2018. PMID: 30035852
-
Serine hydroxymethyltransferase 2: a novel target for human cancer therapy.Invest New Drugs. 2021 Dec;39(6):1671-1681. doi: 10.1007/s10637-021-01144-z. Epub 2021 Jul 3. Invest New Drugs. 2021. PMID: 34215932 Review.
-
Roles of Mitochondrial Serine Hydroxymethyltransferase 2 (SHMT2) in Human Carcinogenesis.J Cancer. 2021 Aug 8;12(19):5888-5894. doi: 10.7150/jca.60170. eCollection 2021. J Cancer. 2021. PMID: 34476002 Free PMC article. Review.
Cited by
-
Unraveling the role of the mitochondrial one-carbon pathway in undifferentiated thyroid cancer by multi-omics analyses.Nat Commun. 2024 Feb 8;15(1):1163. doi: 10.1038/s41467-024-45366-0. Nat Commun. 2024. PMID: 38331894 Free PMC article.
-
The RNA-binding protein landscapes differ between mammalian organs and cultured cells.Nat Commun. 2023 Apr 12;14(1):2074. doi: 10.1038/s41467-023-37494-w. Nat Commun. 2023. PMID: 37045843 Free PMC article.
-
Targeting HOTAIRM1 ameliorates glioblastoma by disrupting mitochondrial oxidative phosphorylation and serine metabolism.iScience. 2022 Jul 31;25(8):104823. doi: 10.1016/j.isci.2022.104823. eCollection 2022 Aug 19. iScience. 2022. PMID: 35992092 Free PMC article.
-
The role of serine metabolism in lung cancer: From oncogenesis to tumor treatment.Front Genet. 2023 Jan 9;13:1084609. doi: 10.3389/fgene.2022.1084609. eCollection 2022. Front Genet. 2023. PMID: 36699468 Free PMC article. Review.
-
RNA-binding proteins in cardiovascular biology and disease: the beat goes on.Nat Rev Cardiol. 2024 Jun;21(6):361-378. doi: 10.1038/s41569-023-00958-z. Epub 2024 Jan 2. Nat Rev Cardiol. 2024. PMID: 38163813 Review.
References
-
- Liu J., Schmitz J.C., Lin X., Tai N., Yan W., Farrell M., Bailly M., Chen T.M., Chu E.. Thymidylate synthase as a translational regulator of cellular gene expression. Biochim. Biophys. Acta. 2002; 1587:174–182. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
