

Humans as inverted bats: A comparative approach to the obstetric conundrum
- PMID: 30810261
- PMCID: PMC6492174
- DOI: 10.1002/ajhb.23227
Humans as inverted bats: A comparative approach to the obstetric conundrum
Abstract
Objectives: The narrow human birth canal evolved in response to multiple opposing selective forces on the pelvis. These factors cannot be sufficiently disentangled in humans because of the limited range of relevant variation. Here, we outline a comparative strategy to study the evolution of human childbirth and to test existing hypotheses in primates and other mammals.
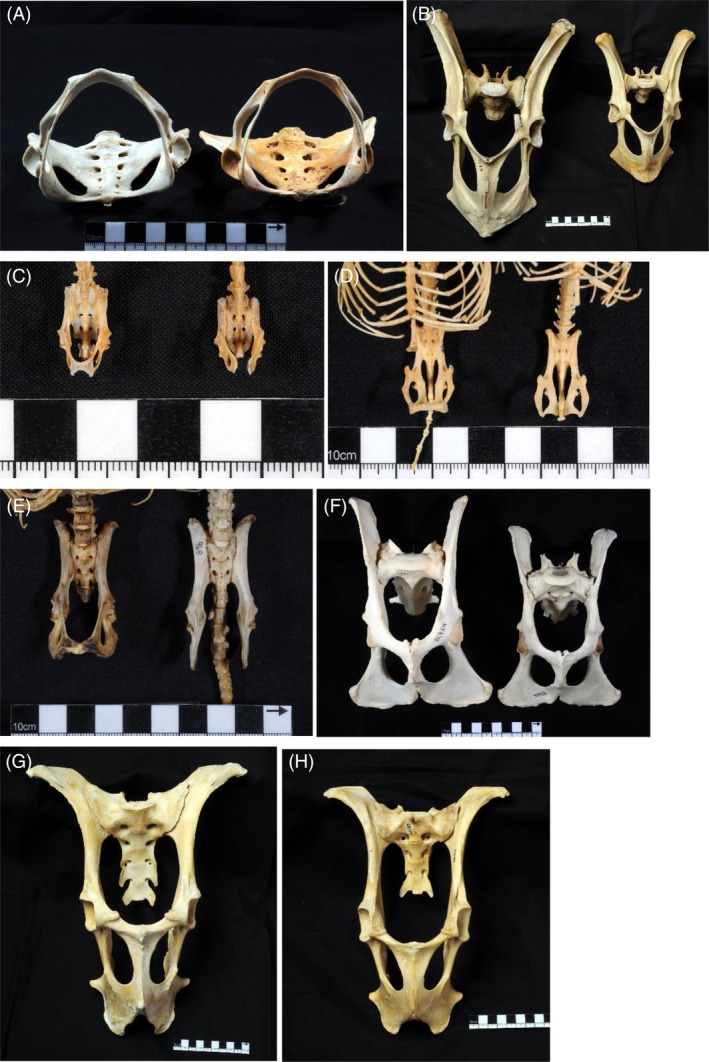
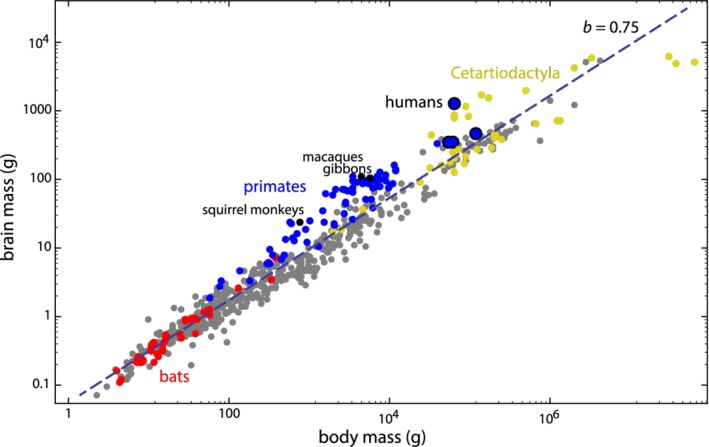
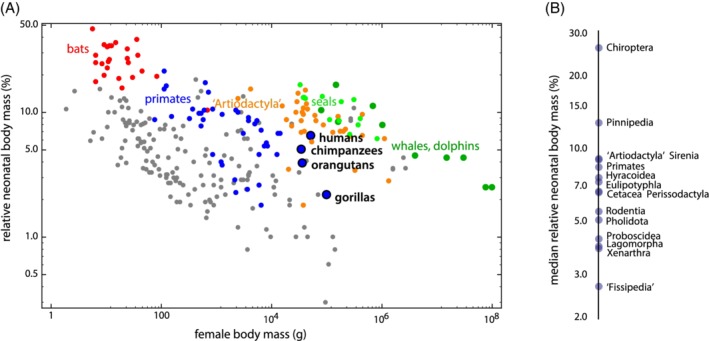
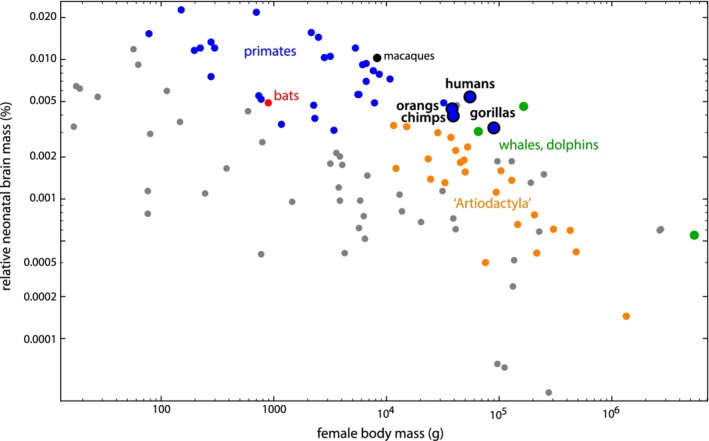
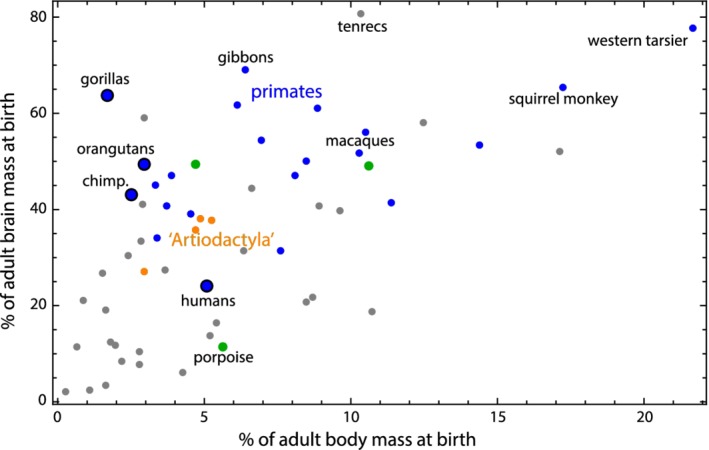
Methods: We combined a literature review with comparative analyses of neonatal and female body and brain mass, using three existing datasets. We also present images of bony pelves of a diverse sample of taxa.
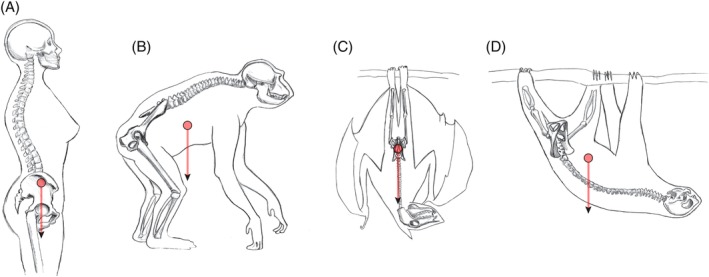
Results: Bats, certain non-human primates, seals, and most ungulates, including whales, have much larger relative neonatal masses than humans, and they all differ in their anatomical adaptations for childbirth. Bats, as a group, are particularly interesting in this context as they give birth to the relatively largest neonates, and their pelvis is highly dimorphic: Whereas males have a fused symphysis, a ligament bridges a large pubic gap in females. The resulting strong demands on the widened and vulnerable pelvic floor likely are relaxed by roosting head-down.
Conclusions: Parturition has constituted a strong selective force in many non-human placentals. We illustrated how the demands on pelvic morphology resulting from locomotion, pelvic floor stability, childbirth, and perhaps also erectile function in males have been traded off differently in mammals, depending on their locomotion and environment. Exploiting the power of a comparative approach, we present new hypotheses and research directions for resolving the obstetric conundrum in humans.
© 2019 The Authors. American Journal of Human Biology published by Wiley Periodicals, Inc.
Figures
References
-
- Abee, C. R. (1989). The squirrel monkey in biomedical research. Institute for Laboratory Animal Research Journal, 31, 11–20.
-
- Abitbol, M. M. (1988). Evolution of the ischial spine of the pelvic floor in the Hominoidea. American Journal of Physical Anthropology, 75, 53–67. - PubMed
-
- Adams, R. A. , & Thibault, K. M. (1999). Growth, development, and histology of the calcar in the little brown bat, Myotis lucifugus (Vespertilionidae). Acta Chiropterologica, 1(2), 215–221.
-
- Aksel, S. , & Abee, C. R. (1983). A pelvimetry method for predicting perinatal mortality in pregnant squirrel monkeys (Saimiri sciureus) . Laboratory Animal Science, 33(2), 165–167. - PubMed
-
- Aldridge, H. , & Rautenbach, I. (1987). Morphology, echolocation and resource partitioning in insectivorous bats. The Journal of Animal Ecology, 56(3), 763–778.
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
