

Early splicing functions of fission yeast Prp16 and its unexpected requirement for gene Silencing is governed by intronic features
- PMID: 30810475
- PMCID: PMC6546409
- DOI: 10.1080/15476286.2019.1585737
Early splicing functions of fission yeast Prp16 and its unexpected requirement for gene Silencing is governed by intronic features
Abstract
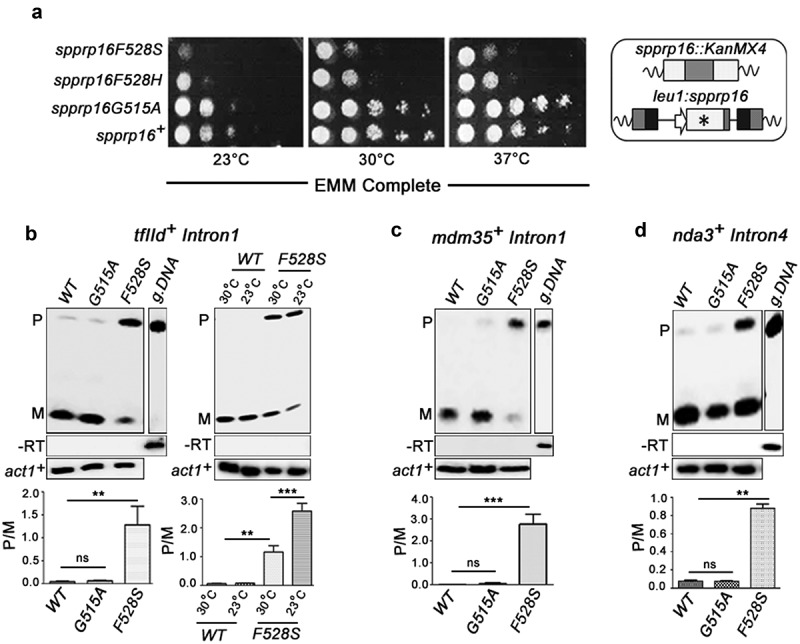
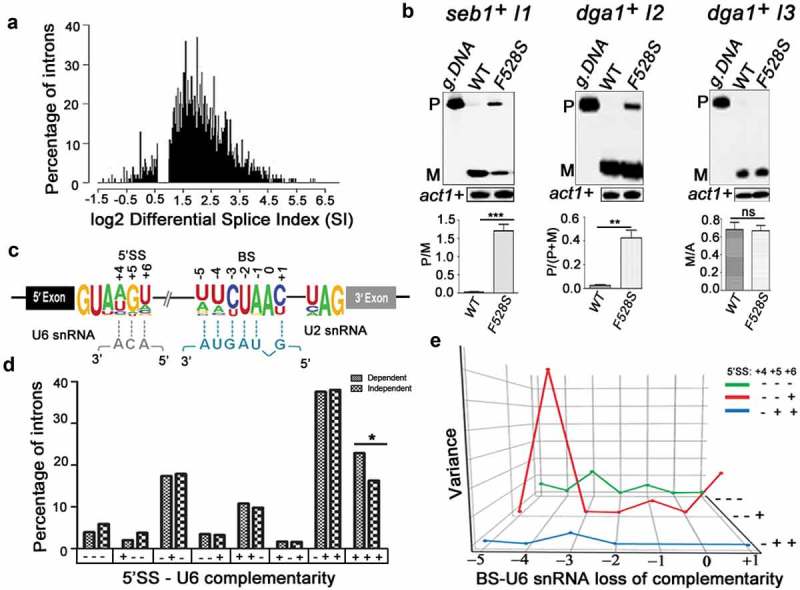
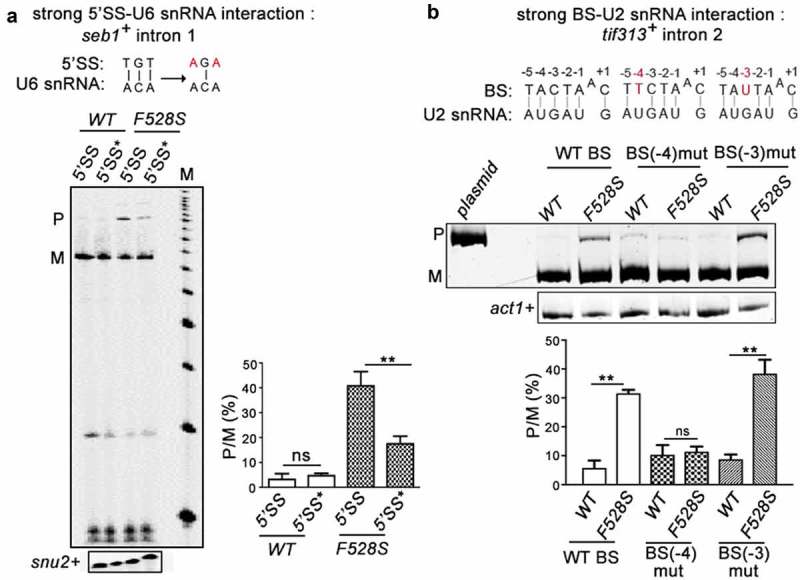
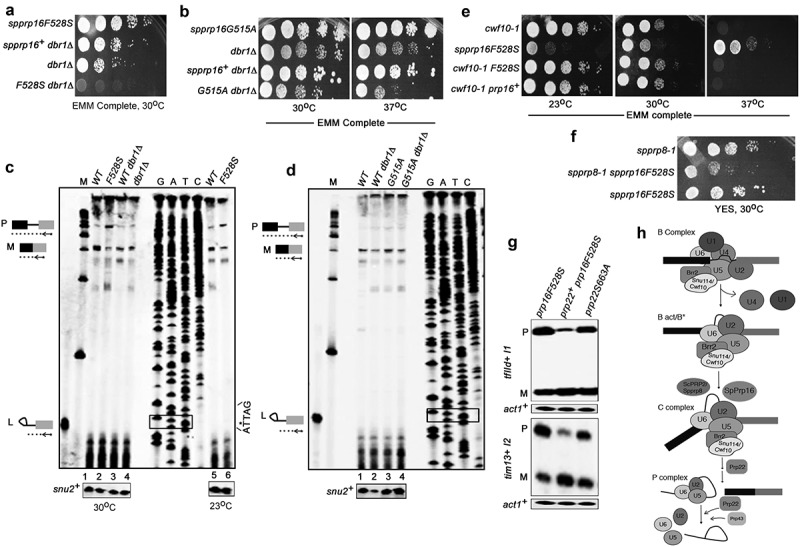
Prp16 is a DEAH box pre-mRNA splicing factor that triggers a key spliceosome conformational switch to facilitate second step splicing in Saccharomyces cerevisiae. However, Prp16 functions are largely unexplored in Schizosaccharomyces pombe, an attractive model with exon-intron architecture more relevant to several other eukaryotes. Here, we generated mis-sense alleles in SpPrp16 whose consequences on genome-wide splicing uncover its nearly global splicing role with only a small subset of unaffected introns. Prp16 dependent and independent intron categories displayed a striking difference in the strength of intronic 5' splice site (5'SS)-U6 snRNA and branch site (BS)-U2 snRNA interactions. Selective weakening of these interactions could convert a Prp16 dependent intron into an independent one. These results point to the role of SpPrp16 in destabilizing 5'SS-U6snRNA and BS-U2snRNA interactions which plausibly trigger structural alterations in the spliceosome to facilitate first step catalysis. Our data suggest that SpPrp16 interactions with early acting factors, its enzymatic activities and association with intronic elements collectively account for efficient and accurate first step catalysis. In addition to splicing derangements in the spprp16F528S mutant, we show that SpPrp16 influences cell cycle progression and centromeric heterochromatinization. We propose that strong 5'SS-U6 snRNA and BS-U2 snRNA complementarity of intron-like elements in non-coding RNAs which lead to complete splicing arrest and impaired Seb1 functions at the pericentromeric loci may cumulatively account for the heterochromatin defects in spprp16F528S cells. These findings suggest that the diverse Prp16 functions within a genome are likely governed by its intronic features that influence splice site-snRNA interaction strength.
Keywords: SpPrp16; cell-cycle; heterochromatin; interaction; intron-like; ncRNA; snRNA; splice-site; transcriptome.
Figures
Similar articles
-
Remodeling of U2-U6 snRNA helix I during pre-mRNA splicing by Prp16 and the NineTeen Complex protein Cwc2.Nucleic Acids Res. 2014 Jul;42(12):8008-23. doi: 10.1093/nar/gku431. Epub 2014 May 21. Nucleic Acids Res. 2014. PMID: 24848011 Free PMC article.
-
Genetic interactions between the yeast RNA helicase homolog Prp16 and spliceosomal snRNAs identify candidate ligands for the Prp16 RNA-dependent ATPase.Genetics. 1994 Jul;137(3):677-87. doi: 10.1093/genetics/137.3.677. Genetics. 1994. PMID: 8088513 Free PMC article.
-
Characterization of Cwc2, U6 snRNA, and Prp8 interactions destabilized by Prp16 ATPase at the transition between the first and second steps of splicing.RNA. 2024 Aug 16;30(9):1199-1212. doi: 10.1261/rna.079886.123. RNA. 2024. PMID: 38876504 Free PMC article.
-
RNA Splicing by the Spliceosome.Annu Rev Biochem. 2020 Jun 20;89:359-388. doi: 10.1146/annurev-biochem-091719-064225. Epub 2019 Dec 3. Annu Rev Biochem. 2020. PMID: 31794245 Review.
-
Recognition of the 5' splice site by the spliceosome.Acta Biochim Pol. 1998;45(4):869-81. Acta Biochim Pol. 1998. PMID: 10397335 Review.
Cited by
-
Centromeric Transcription: A Conserved Swiss-Army Knife.Genes (Basel). 2020 Aug 9;11(8):911. doi: 10.3390/genes11080911. Genes (Basel). 2020. PMID: 32784923 Free PMC article. Review.
-
Prp16 enables efficient splicing of introns with diverse exonic consensus elements in the short-intron rich Cryptococcus neoformans transcriptome.RNA Biol. 2025 Dec;22(1):1-14. doi: 10.1080/15476286.2025.2477844. Epub 2025 Mar 13. RNA Biol. 2025. PMID: 40065603 Free PMC article.
-
Knockout of dhx38 Causes Inner Ear Developmental Defects in Zebrafish.Biomedicines. 2024 Dec 26;13(1):20. doi: 10.3390/biomedicines13010020. Biomedicines. 2024. PMID: 39857604 Free PMC article.
-
Centromeric and pericentric transcription and transcripts: their intricate relationships, regulation, and functions.Chromosoma. 2023 Sep;132(3):211-230. doi: 10.1007/s00412-023-00801-x. Epub 2023 Jul 4. Chromosoma. 2023. PMID: 37401943 Free PMC article. Review.
-
Cryptococcus neoformans Slu7 ensures nuclear positioning during mitotic progression through RNA splicing.PLoS Genet. 2024 May 20;20(5):e1011272. doi: 10.1371/journal.pgen.1011272. eCollection 2024 May. PLoS Genet. 2024. PMID: 38768219 Free PMC article.
References
-
- Wahl MC, Will CL, Luhrmann R.. The spliceosome: design principles of a dynamic RNP machine. Cell. 2009. February 20;136(4):701–718. PubMed PMID: 19239890; Eng. - PubMed
-
- Staley JP, Guthrie C. Mechanical devices of the spliceosome: motors, clocks, springs, and things. Cell. 1998. February 6;92(3):315–326. PubMed PMID: 9476892; eng. - PubMed
-
- Schwer B. A new twist on RNA helicases: DExH/D box proteins as RNPases. Nat Struct Biol. 2001. February;8(2):113–116. PubMed PMID: 11175897; Eng. - PubMed
-
- Chen JY, Stands L, Staley JP, et al. Specific alterations of U1-C protein or U1 small nuclear RNA can eliminate the requirement of Prp28p, an essential DEAD box splicing factor. Mol Cell. 2001. January;7(1):227–232. PubMed PMID: 11172727; eng. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases