

Endothelin receptor Aa regulates proliferation and differentiation of Erb-dependent pigment progenitors in zebrafish
- PMID: 30811380
- PMCID: PMC6392274
- DOI: 10.1371/journal.pgen.1007941
Endothelin receptor Aa regulates proliferation and differentiation of Erb-dependent pigment progenitors in zebrafish
Abstract
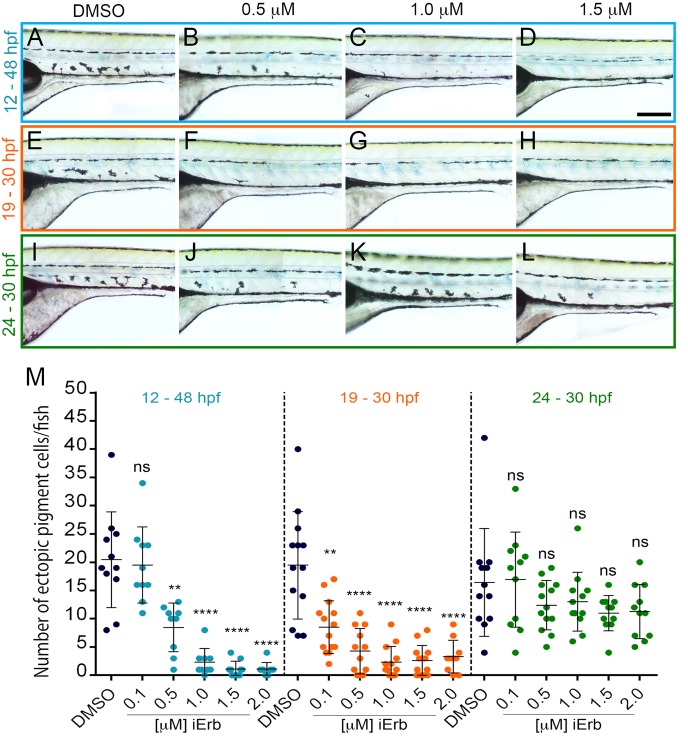
Skin pigment patterns are important, being under strong selection for multiple roles including camouflage and UV protection. Pigment cells underlying these patterns form from adult pigment stem cells (APSCs). In zebrafish, APSCs derive from embryonic neural crest cells, but sit dormant until activated to produce pigment cells during metamorphosis. The APSCs are set-aside in an ErbB signaling dependent manner, but the mechanism maintaining quiescence until metamorphosis remains unknown. Mutants for a pigment pattern gene, parade, exhibit ectopic pigment cells localised to the ventral trunk, but also supernumerary cells restricted to the Ventral Stripe. Contrary to expectations, these melanocytes and iridophores are discrete cells, but closely apposed. We show that parade encodes Endothelin receptor Aa, expressed in the blood vessels, most prominently in the medial blood vessels, consistent with the ventral trunk phenotype. We provide evidence that neuronal fates are not affected in parade mutants, arguing against transdifferentiation of sympathetic neurons to pigment cells. We show that inhibition of BMP signaling prevents specification of sympathetic neurons, indicating conservation of this molecular mechanism with chick and mouse. However, inhibition of sympathetic neuron differentiation does not enhance the parade phenotype. Instead, we pinpoint ventral trunk-restricted proliferation of neural crest cells as an early feature of the parade phenotype. Importantly, using a chemical genetic screen for rescue of the ectopic pigment cell phenotype of parade mutants (whilst leaving the embryonic pattern untouched), we identify ErbB inhibitors as a key hit. The time-window of sensitivity to these inhibitors mirrors precisely the window defined previously as crucial for the setting aside of APSCs in the embryo, strongly implicating adult pigment stem cells as the source of the ectopic pigment cells. We propose that a novel population of APSCs exists in association with medial blood vessels, and that their quiescence is dependent upon Endothelin-dependent factors expressed by the blood vessels.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
- MC_UU_00007/9/MRC_/Medical Research Council/United Kingdom
- MC_PC_U127585840/MRC_/Medical Research Council/United Kingdom
- MC_U127585840/MRC_/Medical Research Council/United Kingdom
- BB/L00769X/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- MR/J001457/1/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
