

A Developmental Perspective on Paragangliar Tumorigenesis
- PMID: 30813557
- PMCID: PMC6468609
- DOI: 10.3390/cancers11030273
A Developmental Perspective on Paragangliar Tumorigenesis
Abstract
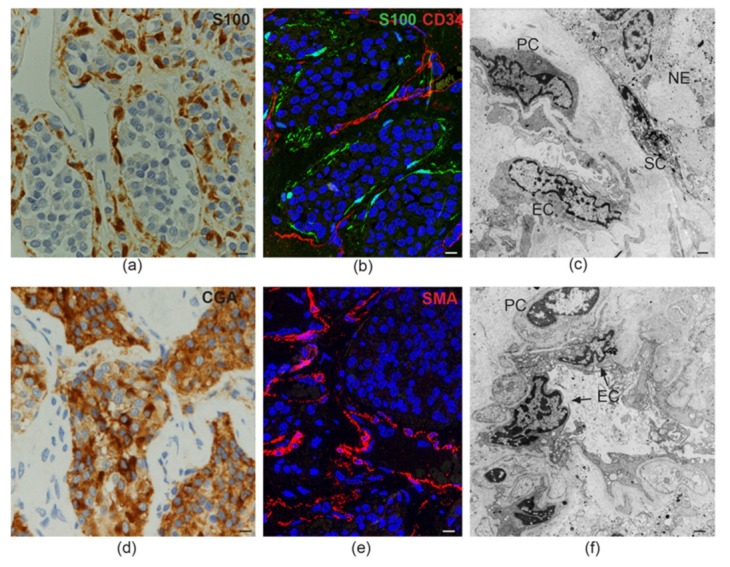
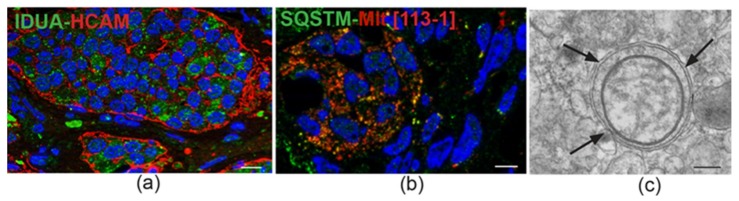
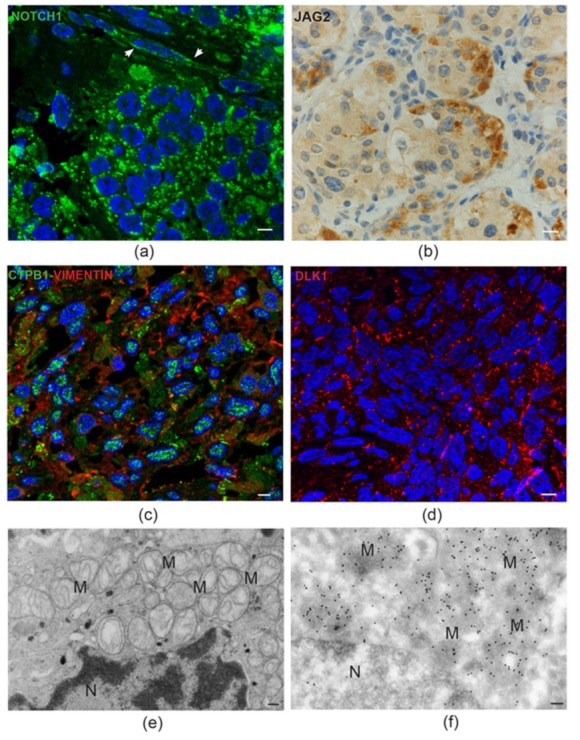
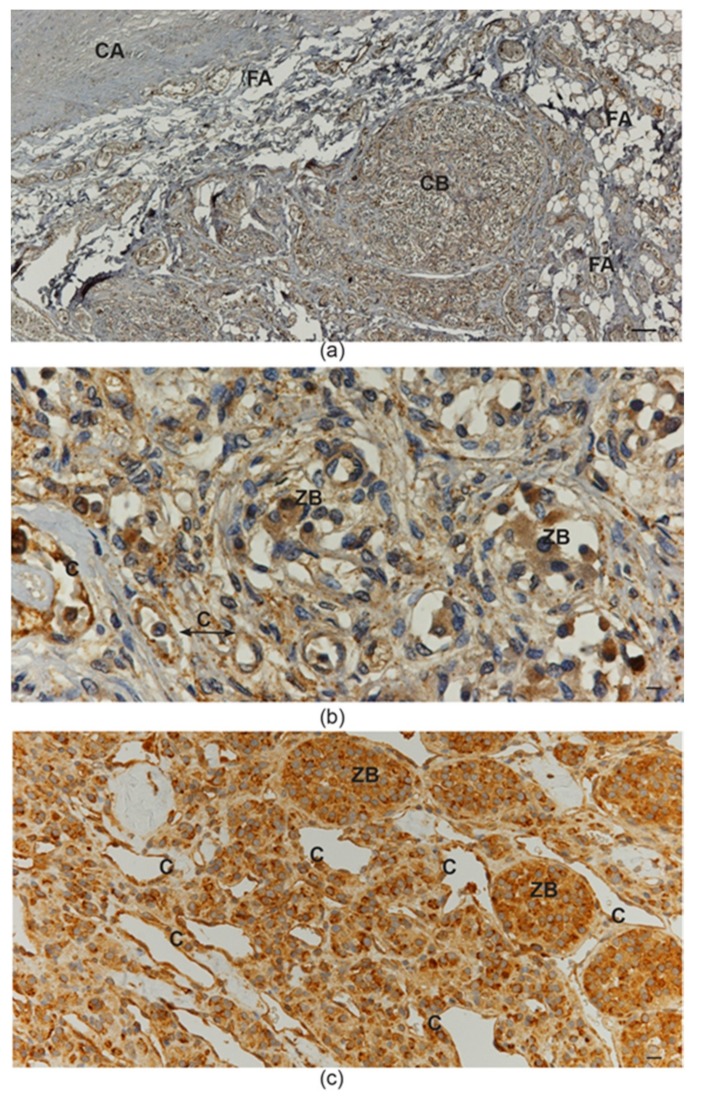
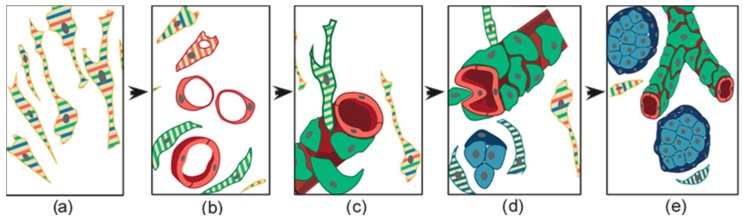
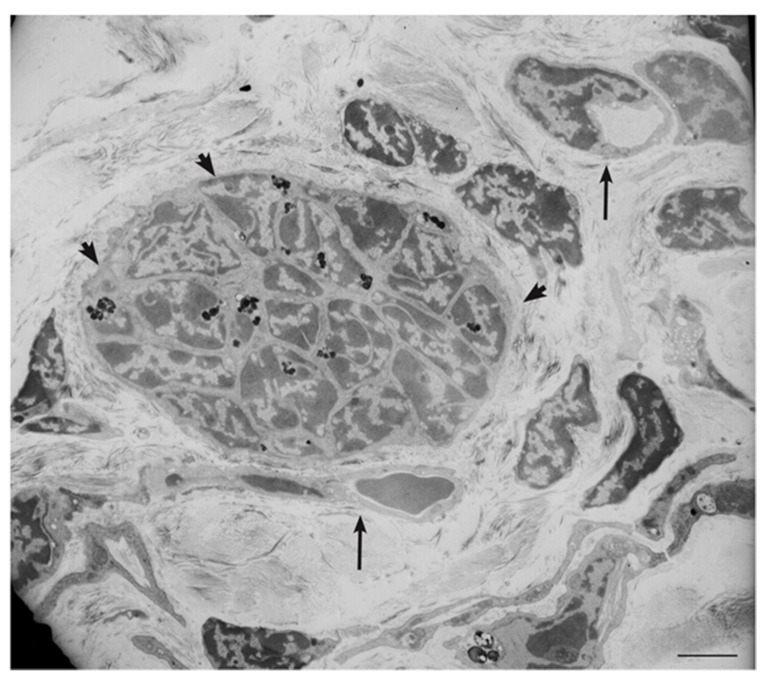
In this review, we propose that paraganglioma is a fundamentally organized, albeit aberrant, tissue composed of neoplastic vascular and neural cell types that share a common origin from a multipotent mesenchymal-like stem/progenitor cell. This view is consistent with the pseudohypoxic footprint implicated in the molecular pathogenesis of the disease, is in harmony with the neural crest origin of the paraganglia, and is strongly supported by the physiological model of carotid body hyperplasia. Our immunomorphological and molecular studies of head and neck paragangliomas demonstrate in all cases relationships between the vascular and the neural tumor compartments, that share mesenchymal and immature vasculo-neural markers, conserved in derived cell cultures. This immature, multipotent phenotype is supported by constitutive amplification of NOTCH signaling genes and by loss of the microRNA-200s and -34s, which control NOTCH1, ZEB1, and PDGFRA in head and neck paraganglioma cells. Importantly, the neuroepithelial component is distinguished by extreme mitochondrial alterations, associated with collapse of the ΔΨm. Finally, our xenograft models of head and neck paraganglioma demonstrate that mesenchymal-like cells first give rise to a vasculo-angiogenic network, and then self-organize into neuroepithelial-like clusters, a process inhibited by treatment with imatinib.
Keywords: angiogenesis; carotid body; mitochondria; neural crest; neurogenesis; paraganglioma; stem-like tumor cells; vasculogenesis; xenograft.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Paragangliomas arise through an autonomous vasculo-angio-neurogenic program inhibited by imatinib.Acta Neuropathol. 2018 May;135(5):779-798. doi: 10.1007/s00401-017-1799-2. Epub 2018 Jan 5. Acta Neuropathol. 2018. PMID: 29305721 Free PMC article.
-
Integrative genetic, epigenetic and pathological analysis of paraganglioma reveals complex dysregulation of NOTCH signaling.Acta Neuropathol. 2013 Oct;126(4):575-94. doi: 10.1007/s00401-013-1165-y. Epub 2013 Aug 18. Acta Neuropathol. 2013. PMID: 23955600 Free PMC article.
-
Basic fibroblast growth factor and fibroblastic growth factor receptor-1 may contribute to head and neck paraganglioma development by an autocrine or paracrine mechanism.Hum Pathol. 2007 Jan;38(1):79-85. doi: 10.1016/j.humpath.2006.06.013. Epub 2006 Aug 1. Hum Pathol. 2007. PMID: 16949906
-
[Phenotypic plasticity of neural crest-derived melanocytes and Schwann cells].Biol Aujourdhui. 2011;205(1):53-61. doi: 10.1051/jbio/2011008. Epub 2011 Apr 19. Biol Aujourdhui. 2011. PMID: 21501576 Review. French.
-
Treatment of Head and Neck Paragangliomas.Cancer Control. 2016 Jul;23(3):228-41. doi: 10.1177/107327481602300306. Cancer Control. 2016. PMID: 27556663 Review.
Cited by
-
The SDHB Arg230His mutation causing familial paraganglioma alters glycolysis in a new Caenorhabditis elegans model.Dis Model Mech. 2020 Oct 15;13(10):dmm044925. doi: 10.1242/dmm.044925. Dis Model Mech. 2020. PMID: 32859697 Free PMC article.
-
Insights into Mechanisms of Pheochromocytomas and Paragangliomas Driven by Known or New Genetic Drivers.Cancers (Basel). 2021 Sep 14;13(18):4602. doi: 10.3390/cancers13184602. Cancers (Basel). 2021. PMID: 34572828 Free PMC article. Review.
-
Pheochromocytoma (PHEO) and Paraganglioma (PGL).Cancers (Basel). 2019 Sep 18;11(9):1391. doi: 10.3390/cancers11091391. Cancers (Basel). 2019. PMID: 31540433 Free PMC article.
-
Targeted metabolomics detects a putatively diagnostic signature in plasma and dried blood spots from head and neck paraganglioma patients.Oncogenesis. 2023 Feb 25;12(1):10. doi: 10.1038/s41389-023-00456-4. Oncogenesis. 2023. PMID: 36841802 Free PMC article.
-
Cancer Stem Cells in Pheochromocytoma and Paraganglioma.Front Endocrinol (Lausanne). 2020 Feb 25;11:79. doi: 10.3389/fendo.2020.00079. eCollection 2020. Front Endocrinol (Lausanne). 2020. PMID: 32158431 Free PMC article. Review.
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
