

Structure-function analyses reveal that a glucuronoyl esterase from Teredinibacter turnerae interacts with carbohydrates and aromatic compounds
- PMID: 30814248
- PMCID: PMC6484129
- DOI: 10.1074/jbc.RA119.007831
Structure-function analyses reveal that a glucuronoyl esterase from Teredinibacter turnerae interacts with carbohydrates and aromatic compounds
Abstract
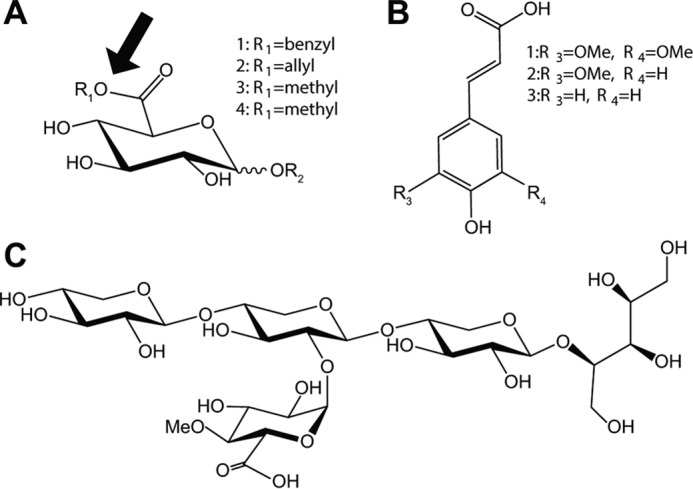
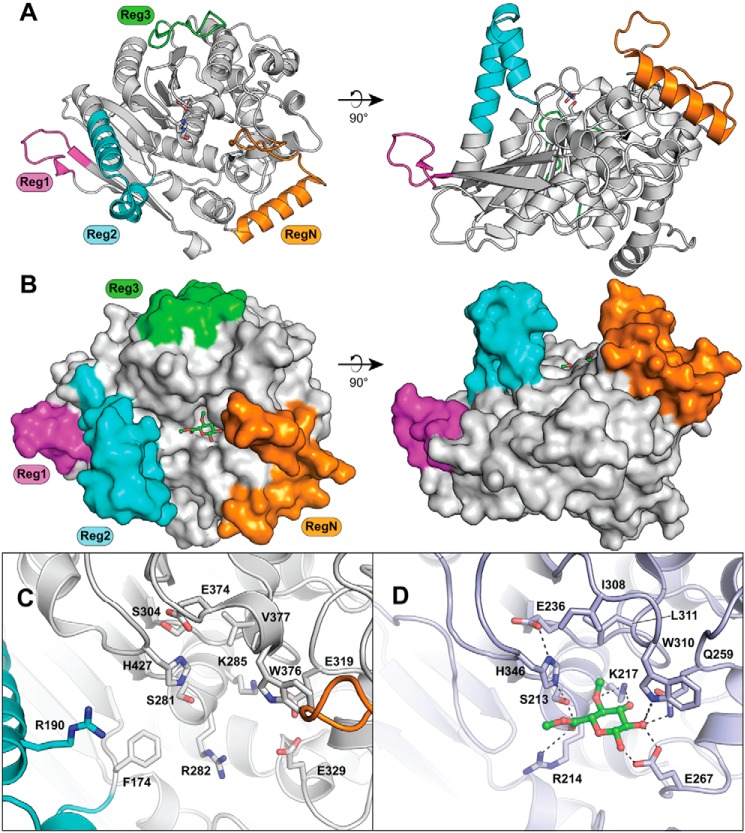
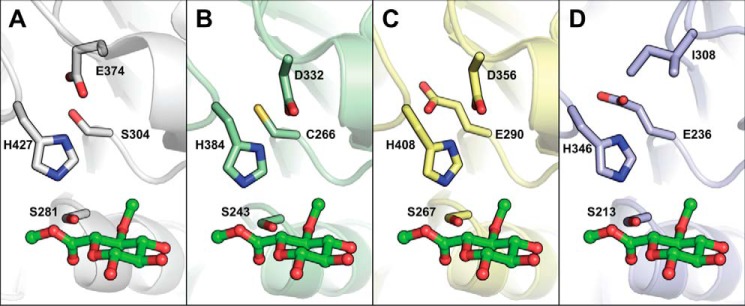
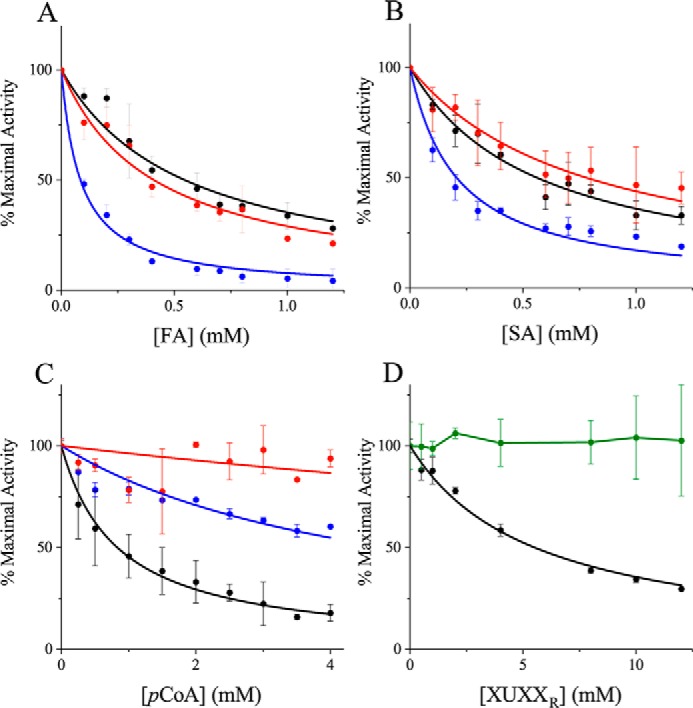
Glucuronoyl esterases (GEs) catalyze the cleavage of ester linkages found between lignin and glucuronic acid moieties on glucuronoxylan in plant biomass. As such, GEs represent promising biochemical tools in industrial processing of these recalcitrant resources. However, details on how GEs interact with their natural substrates are sparse, calling for thorough structure-function studies. Presented here is the structure and biochemical characterization of a GE, TtCE15A, from the bacterium Teredinibacter turnerae, a symbiont of wood-boring shipworms. To gain deeper insight into enzyme-substrate interactions, inhibition studies were performed with both the WT TtCE15A and variants in which we, by using site-directed mutagenesis, substituted residues suggested to have key roles in binding to or interacting with the aromatic and carbohydrate structures of its uronic acid ester substrates. Our results support the hypothesis that two aromatic residues (Phe-174 and Trp-376), conserved in bacterial GEs, interact with aromatic and carbohydrate structures of these substrates in the enzyme active site, respectively. The solved crystal structure of TtCE15A revealed features previously not observed in either fungal or bacterial GEs, with a large inserted N-terminal region neighboring the active site and a differently positioned residue of the catalytic triad. The findings highlight key interactions between GEs and complex lignin-carbohydrate ester substrates and advance our understanding of the substrate specificities of these enzymes in biomass conversion.
Keywords: CE15; biotechnology; carbohydrate; carbohydrate esterase; carbohydrate-active enzymes; enzyme kinetics; enzyme mechanism; enzyme mutation; enzyme structure; glucuronoyl esterase; lignin-carbohydrate complexes; plant cell wall; protein structure; uronic acid.
© 2019 Arnling Bååth et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
