

Circulating myocardial microRNAs from infarcted hearts are carried in exosomes and mobilise bone marrow progenitor cells
- PMID: 30814518
- PMCID: PMC6393447
- DOI: 10.1038/s41467-019-08895-7
Circulating myocardial microRNAs from infarcted hearts are carried in exosomes and mobilise bone marrow progenitor cells
Abstract
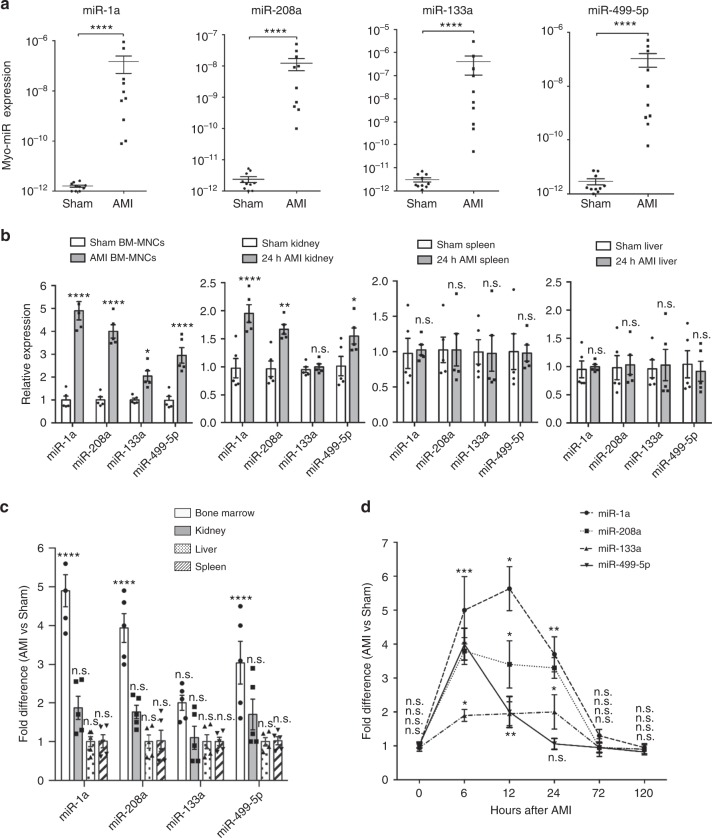
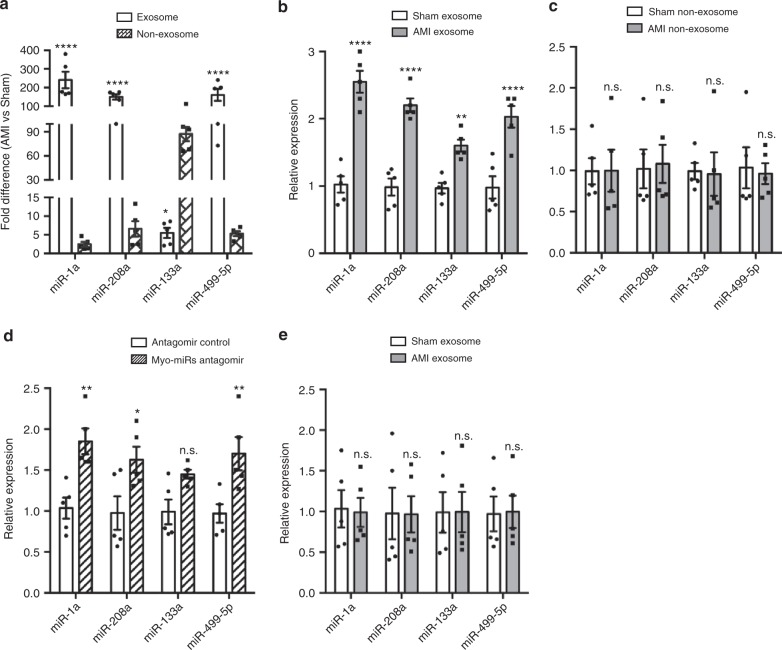
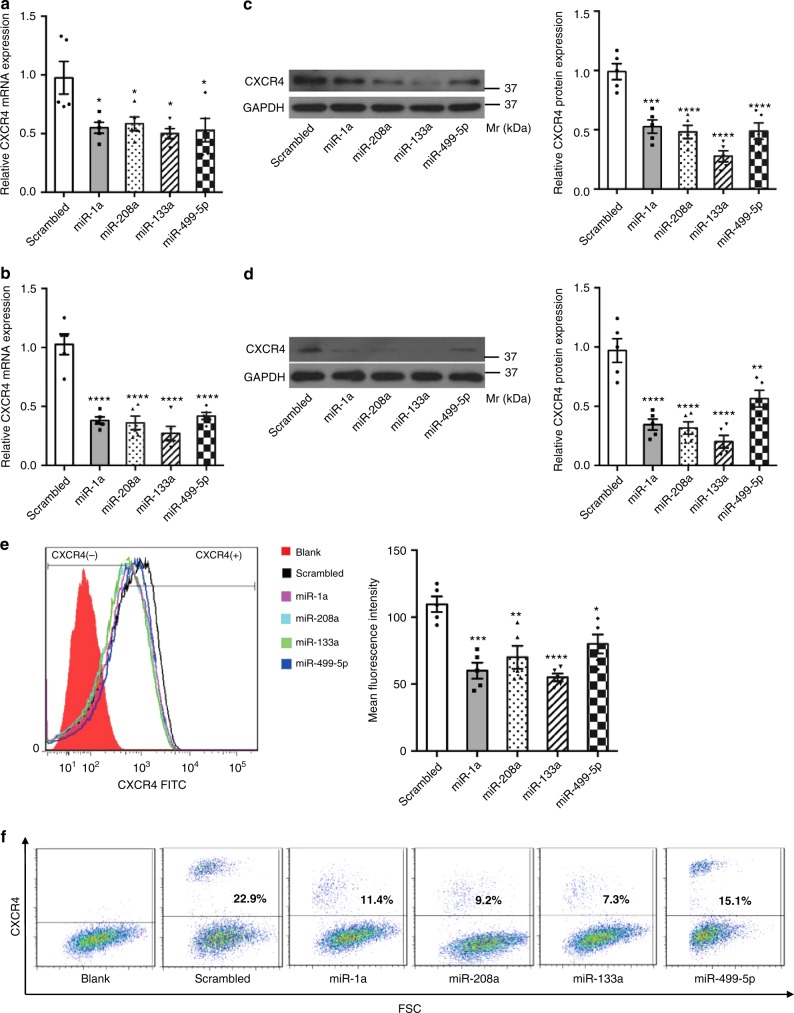
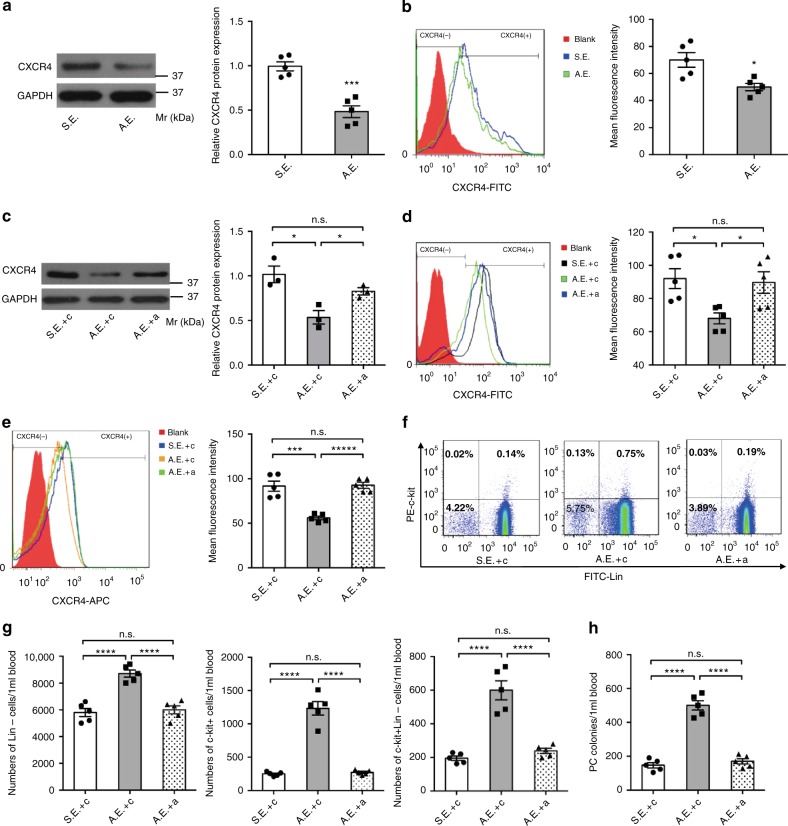
Myocardial microRNAs (myo-miRs) are released into the circulation after acute myocardial infarction (AMI). How they impact remote organs is however largely unknown. Here we show that circulating myo-miRs are carried in exosomes and mediate functional crosstalk between the ischemic heart and the bone marrow (BM). In mice, we find that AMI is accompanied by an increase in circulating levels of myo-miRs, with miR-1, 208, and 499 predominantly in circulating exosomes and miR-133 in the non-exosomal component. Myo-miRs are imported selectively to peripheral organs and preferentially to the BM. Exosomes mediate the transfer of myo-miRs to BM mononuclear cells (MNCs), where myo-miRs downregulate CXCR4 expression. Injection of exosomes isolated from AMI mice into wild-type mice downregulates CXCR4 expression in BM-MNCs and increases the number of circulating progenitor cells. Thus, we propose that myo-miRs carried in circulating exosomes allow a systemic response to cardiac injury that may be leveraged for cardiac repair.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
