

Genome-wide analysis identifies NR4A1 as a key mediator of T cell dysfunction
- PMID: 30814730
- PMCID: PMC6507425
- DOI: 10.1038/s41586-019-0979-8
Genome-wide analysis identifies NR4A1 as a key mediator of T cell dysfunction
Abstract
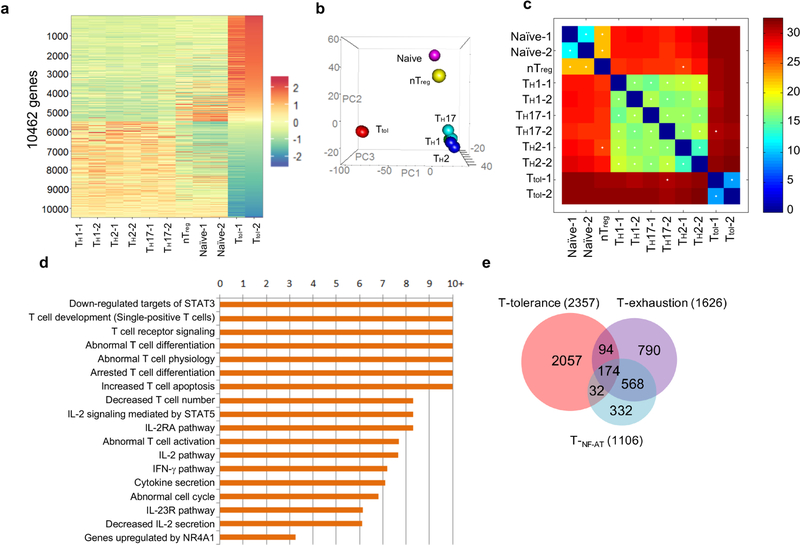
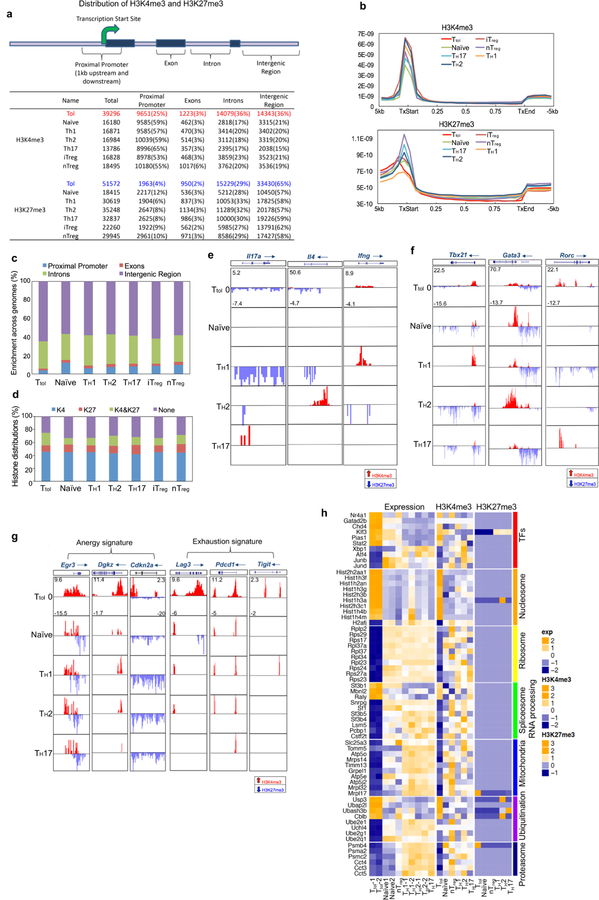
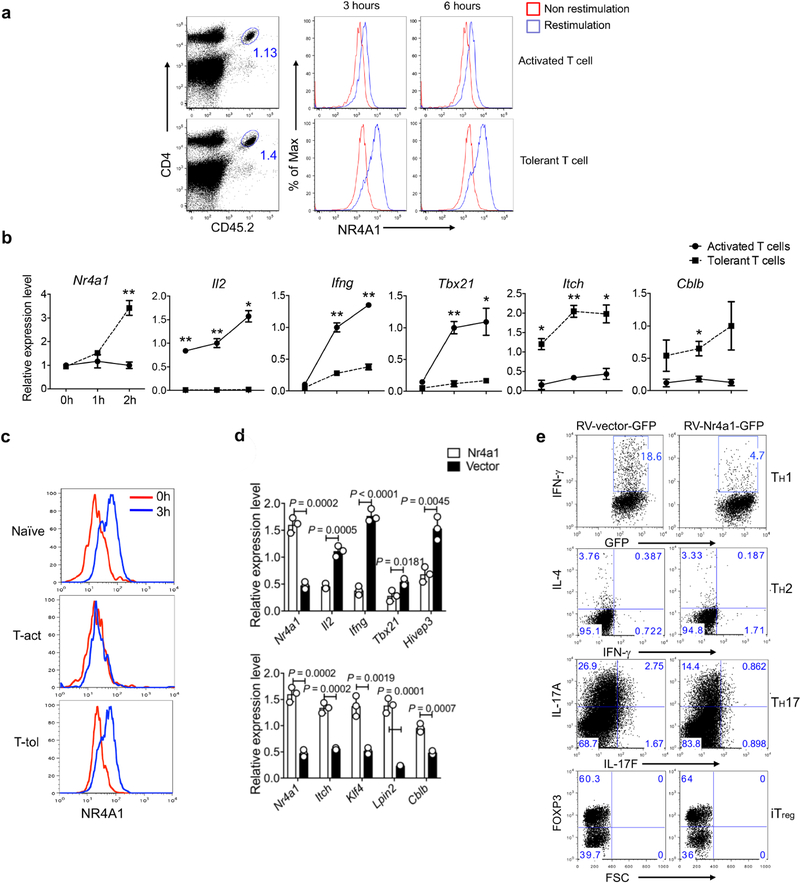
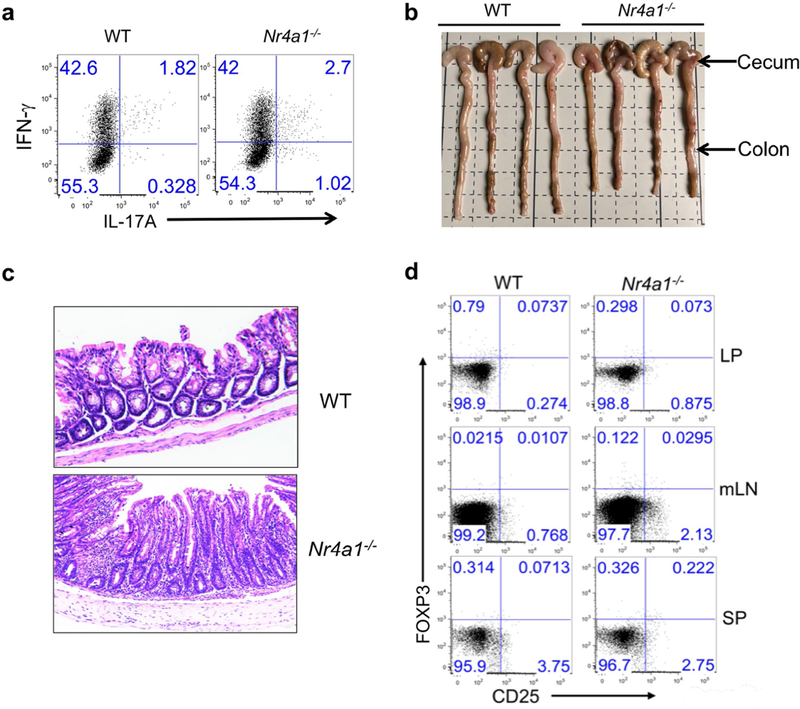
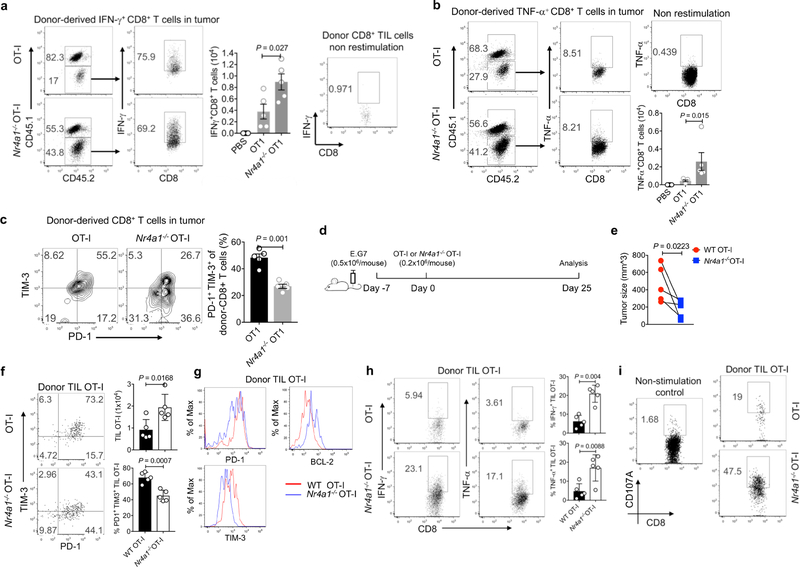
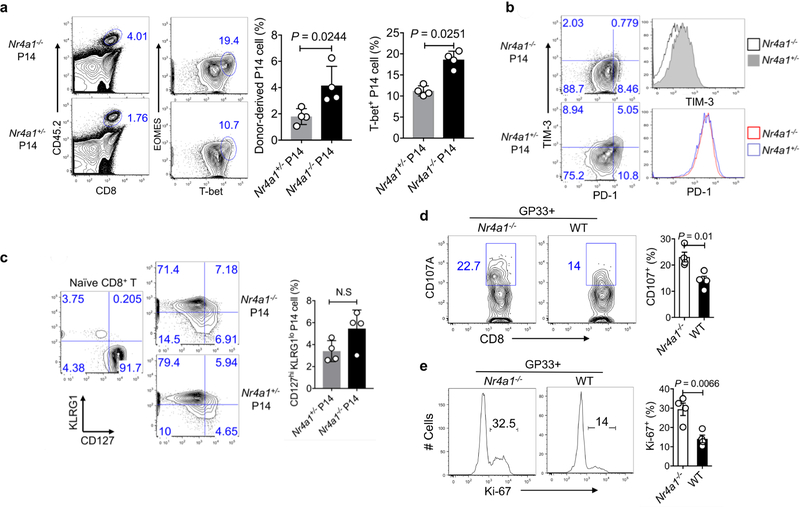
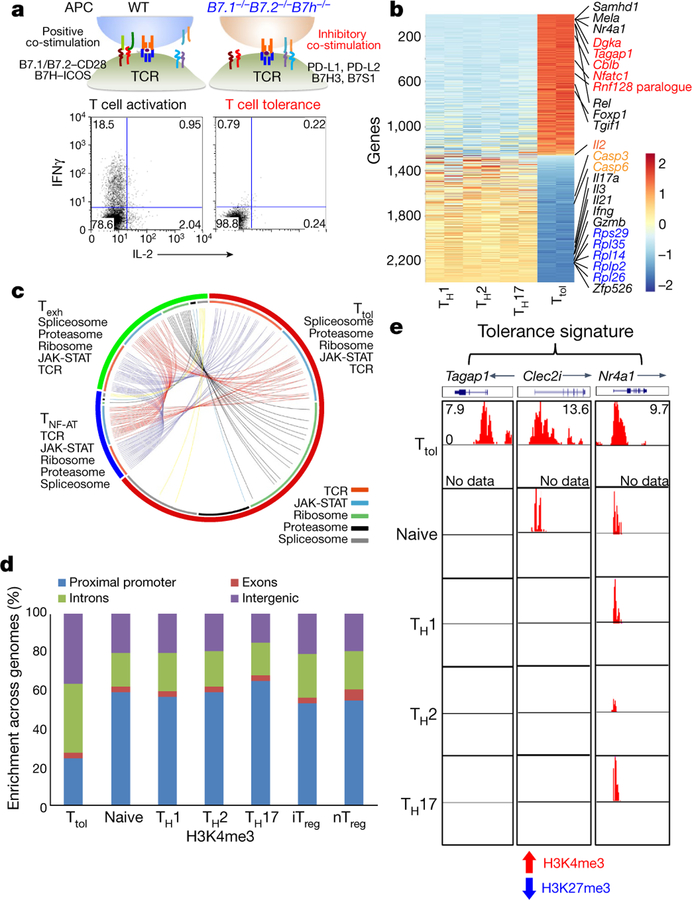
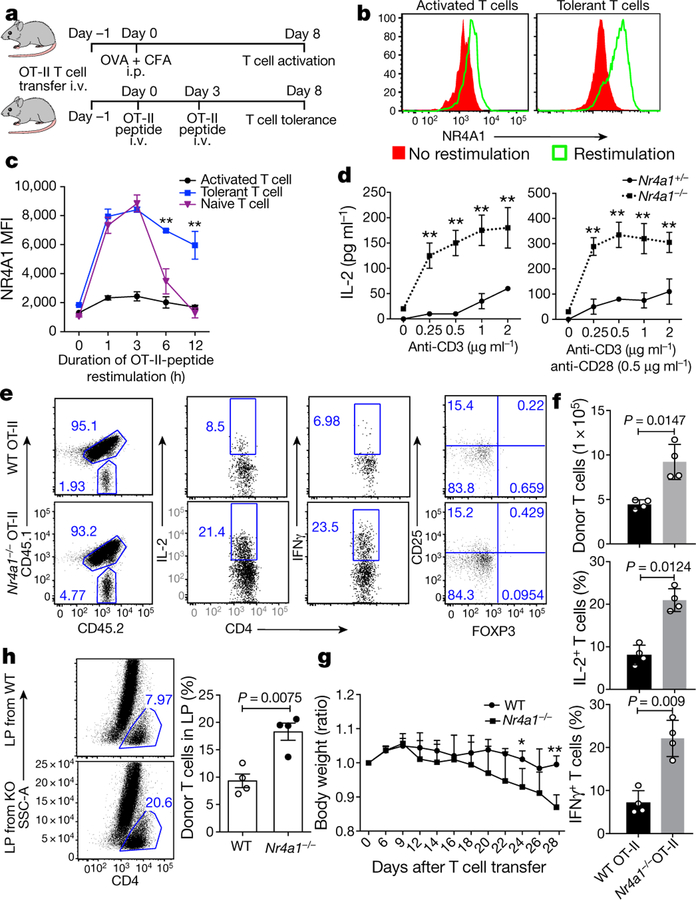
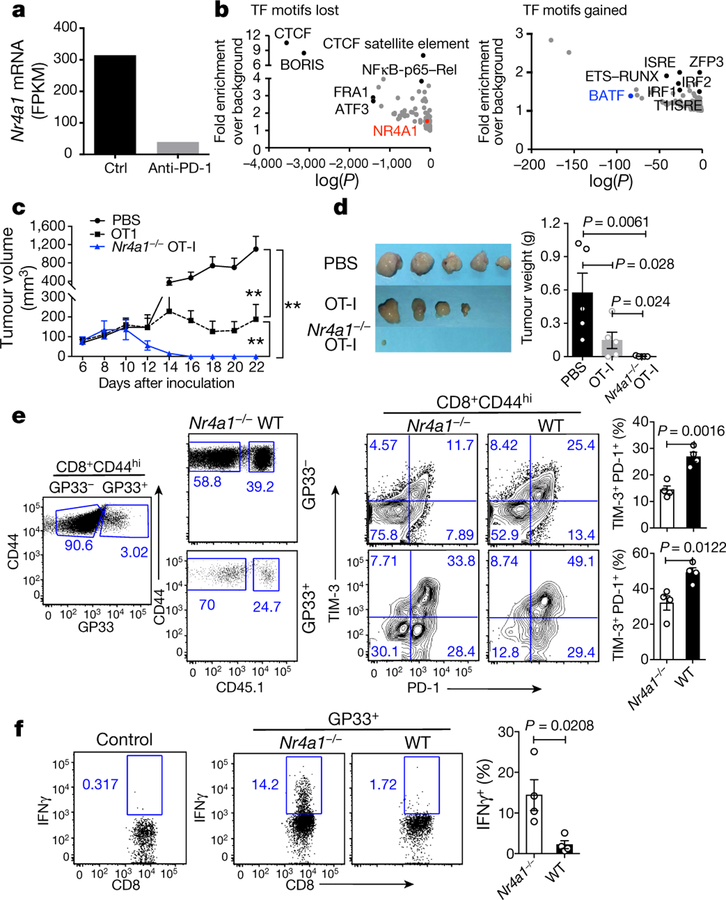
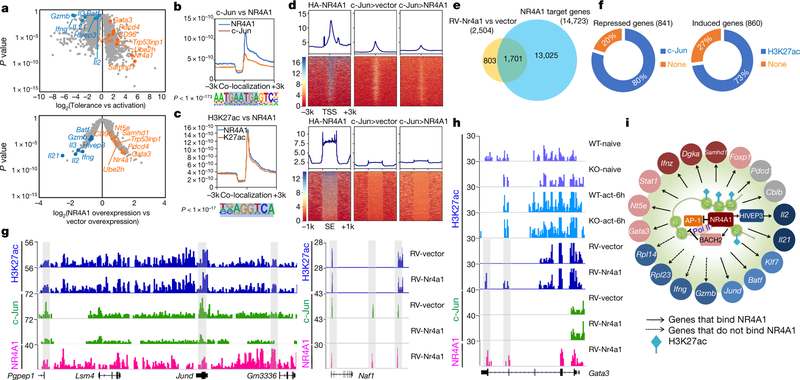
T cells become dysfunctional when they encounter self antigens or are exposed to chronic infection or to the tumour microenvironment1. The function of T cells is tightly regulated by a combinational co-stimulatory signal, and dominance of negative co-stimulation results in T cell dysfunction2. However, the molecular mechanisms that underlie this dysfunction remain unclear. Here, using an in vitro T cell tolerance induction system in mice, we characterize genome-wide epigenetic and gene expression features in tolerant T cells, and show that they are distinct from effector and regulatory T cells. Notably, the transcription factor NR4A1 is stably expressed at high levels in tolerant T cells. Overexpression of NR4A1 inhibits effector T cell differentiation, whereas deletion of NR4A1 overcomes T cell tolerance and exaggerates effector function, as well as enhancing immunity against tumour and chronic virus. Mechanistically, NR4A1 is preferentially recruited to binding sites of the transcription factor AP-1, where it represses effector-gene expression by inhibiting AP-1 function. NR4A1 binding also promotes acetylation of histone 3 at lysine 27 (H3K27ac), leading to activation of tolerance-related genes. This study thus identifies NR4A1 as a key general regulator in the induction of T cell dysfunction, and a potential target for tumour immunotherapy.
Figures



Comment in
-
What's driving T cell dysfunction?Nat Rev Immunol. 2019 Apr;19(4):199. doi: 10.1038/s41577-019-0152-5. Nat Rev Immunol. 2019. PMID: 30858495 No abstract available.
-
What's driving T cell dysfunction?Nat Rev Cancer. 2019 May;19(5):251. doi: 10.1038/s41568-019-0138-4. Nat Rev Cancer. 2019. PMID: 30936467 No abstract available.
References
-
- Greenwald RJ, Freeman GJ & Sharpe AH The B7 family revisited. Annu. Rev. Immunol 23, 515–548 (2005). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
