

Insights into the karyotype and genome evolution of haplogyne spiders indicate a polyploid origin of lineage with holokinetic chromosomes
- PMID: 30816146
- PMCID: PMC6395618
- DOI: 10.1038/s41598-019-39034-3
Insights into the karyotype and genome evolution of haplogyne spiders indicate a polyploid origin of lineage with holokinetic chromosomes
Abstract
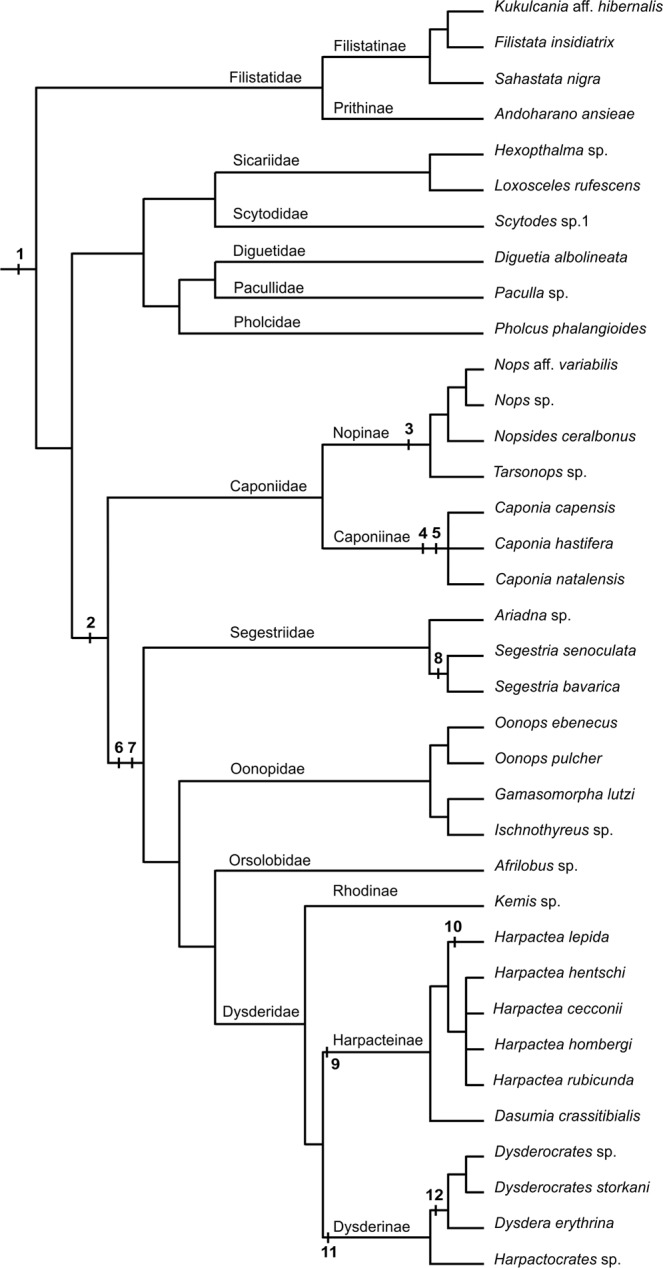
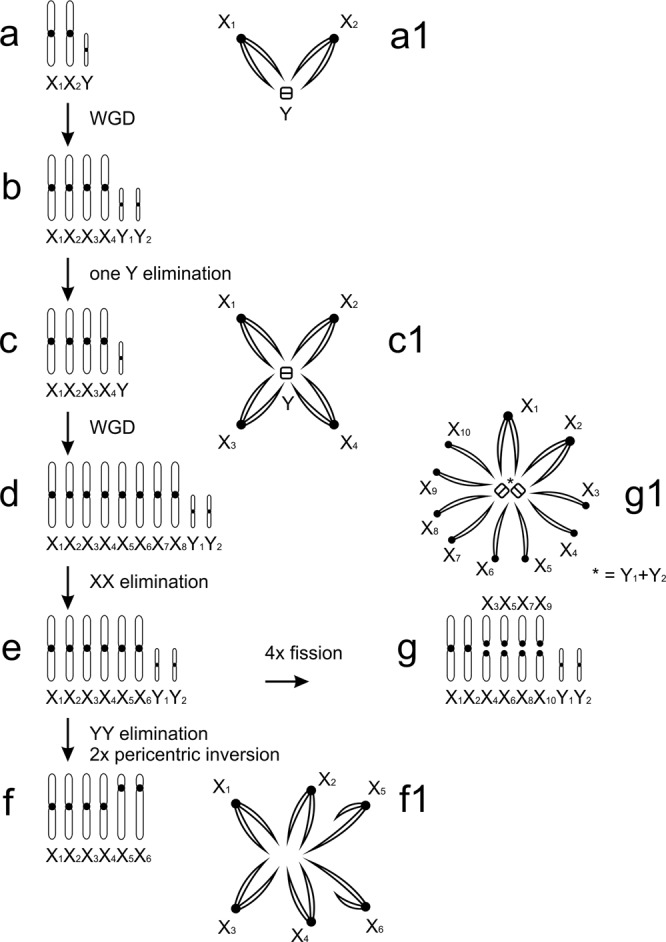
Spiders are an ancient and extremely diverse animal order. They show a considerable diversity of genome sizes, karyotypes and sex chromosomes, which makes them promising models to analyse the evolution of these traits. Our study is focused on the evolution of the genome and chromosomes in haplogyne spiders with holokinetic chromosomes. Although holokinetic chromosomes in spiders were discovered a long time ago, information on their distribution and evolution in these arthropods is very limited. Here we show that holokinetic chromosomes are an autapomorphy of the superfamily Dysderoidea. According to our hypothesis, the karyotype of ancestral Dysderoidea comprised three autosome pairs and a single X chromosome. The subsequent evolution has frequently included inverted meiosis of the sex chromosome and an increase of 2n. We demonstrate that caponiids, a sister clade to Dysderoidea, have enormous genomes and high diploid and sex chromosome numbers. This pattern suggests a polyploid event in the ancestors of caponiids. Holokinetic chromosomes could have arisen by subsequent multiple chromosome fusions and a considerable reduction of the genome size. We propose that spider sex chromosomes probably do not pose a major barrier to polyploidy due to specific mechanisms that promote the integration of sex chromosome copies into the genome.
Conflict of interest statement
The authors declare no competing interests.
Figures
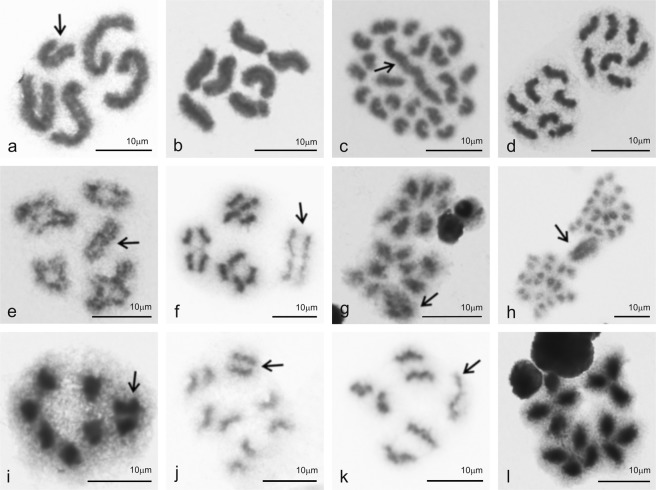
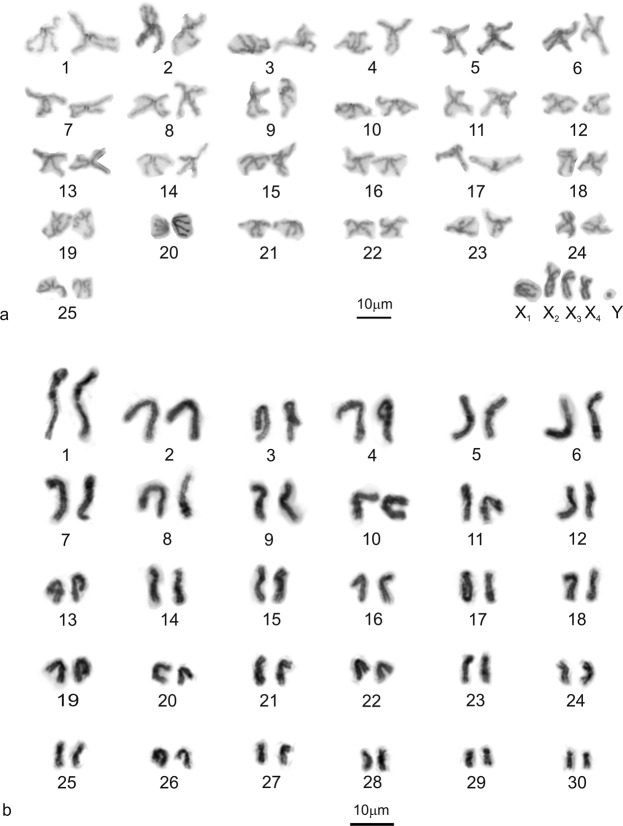
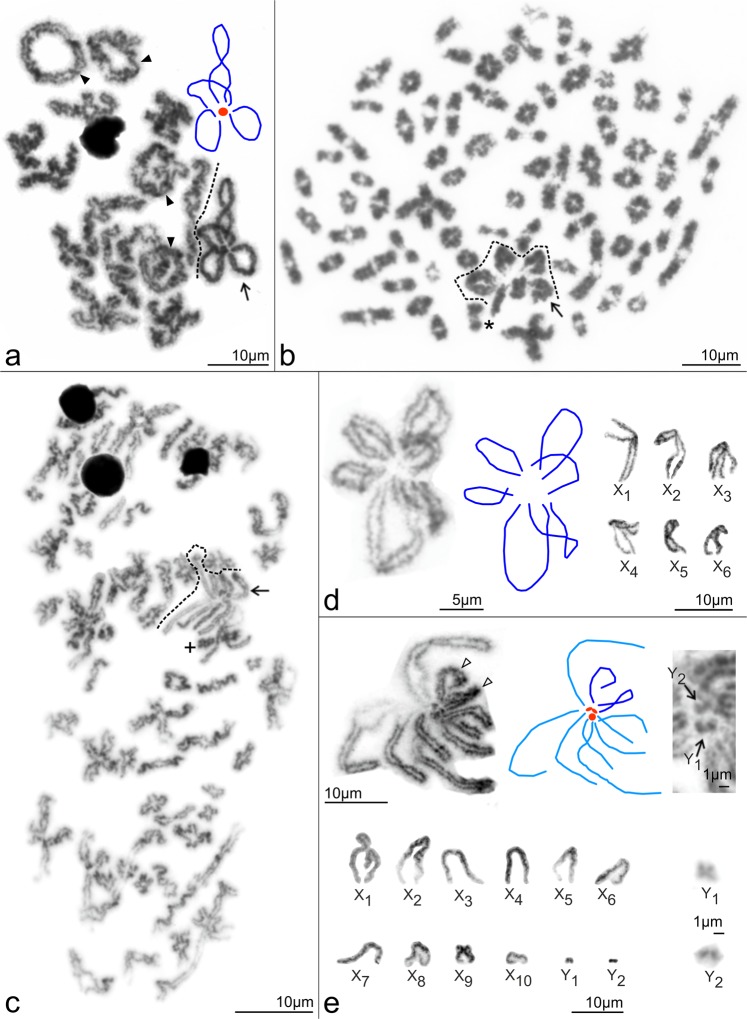
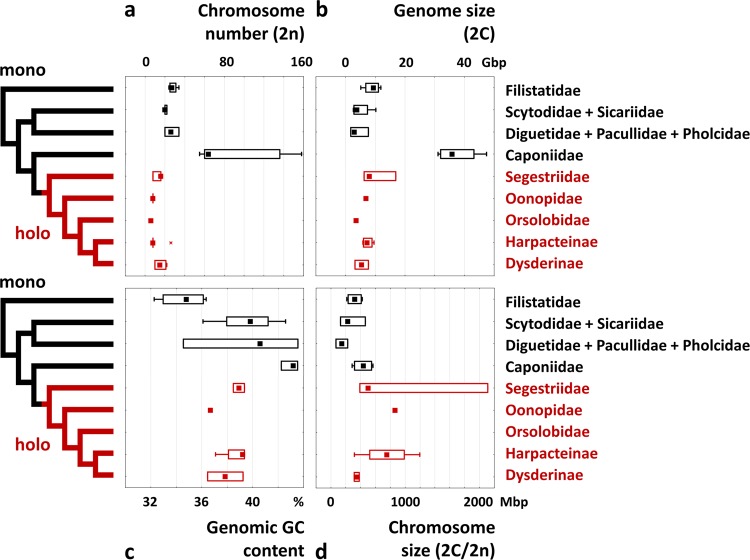
References
-
- Maddison WP. XXXY sex chromosomes in males of the jumping spider genus Pellenes (Araneae: Salticidae) Chromosoma. 1982;5:23–37. doi: 10.1007/BF00344592. - DOI
-
- Rowell DM. Complex sex-linked fusion heterozygosity in the Australian huntsman spider Delena cancerides (Araneae: Sparassidae) Chromosoma. 1985;93:169–176. doi: 10.1007/BF00293165. - DOI
-
- Araujo, D., Schneider, M. C., Paula-Neto, E. & Cella, D. M. Sex chromosomes and meiosis in spiders: a review [Swan, A. (ed.)] Meiosis molecular mechanisms and cytogenetic diversity 87–108 (In Tech, 2012).
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
