

Cold-priming of chloroplast ROS signalling is developmentally regulated and is locally controlled at the thylakoid membrane
- PMID: 30816299
- PMCID: PMC6395587
- DOI: 10.1038/s41598-019-39838-3
Cold-priming of chloroplast ROS signalling is developmentally regulated and is locally controlled at the thylakoid membrane
Abstract
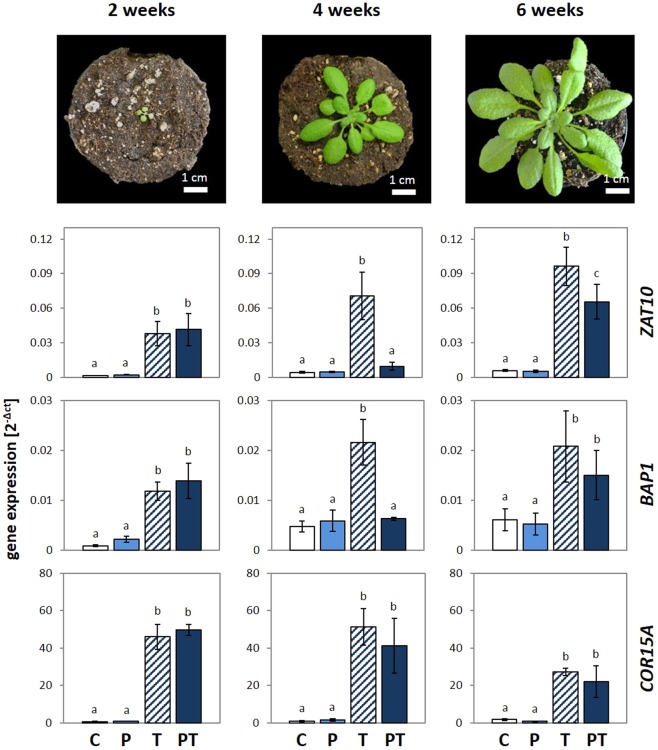
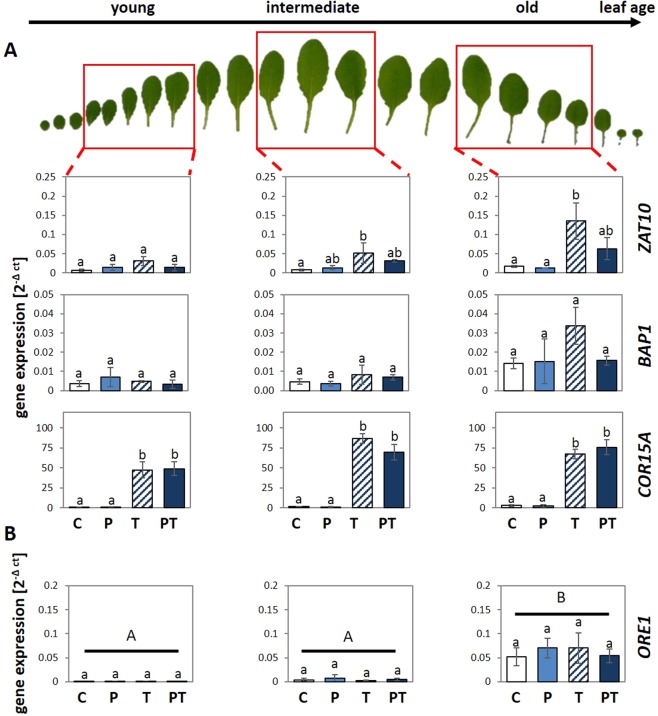
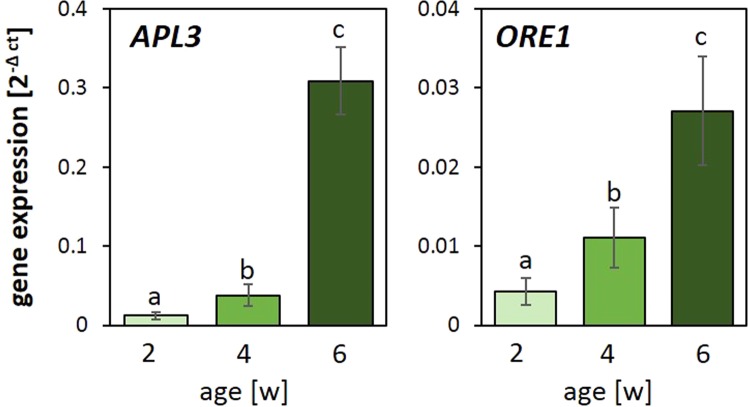
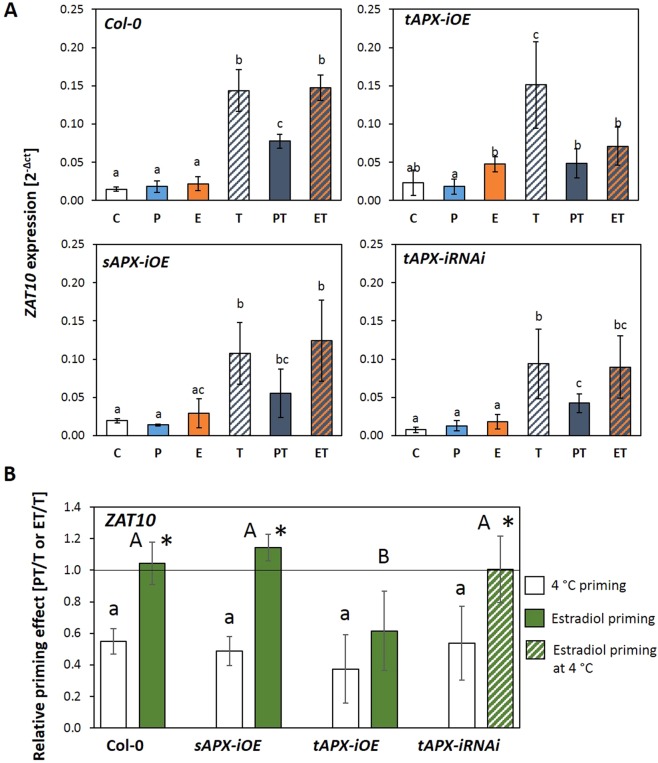
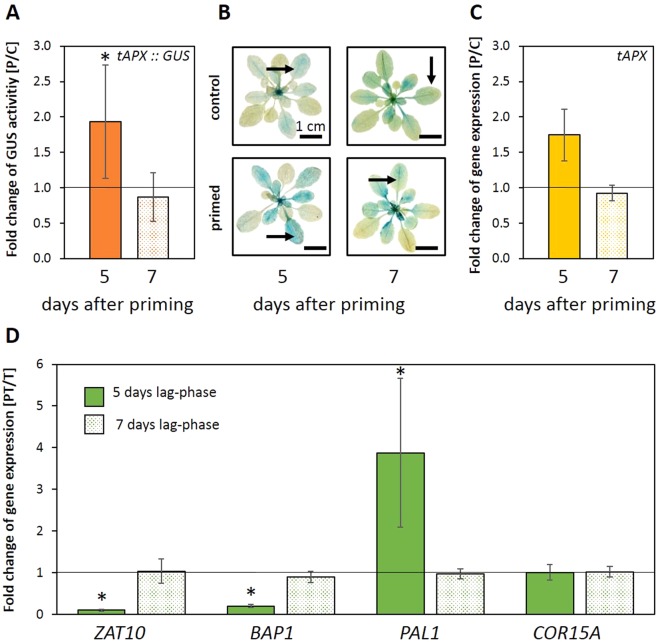
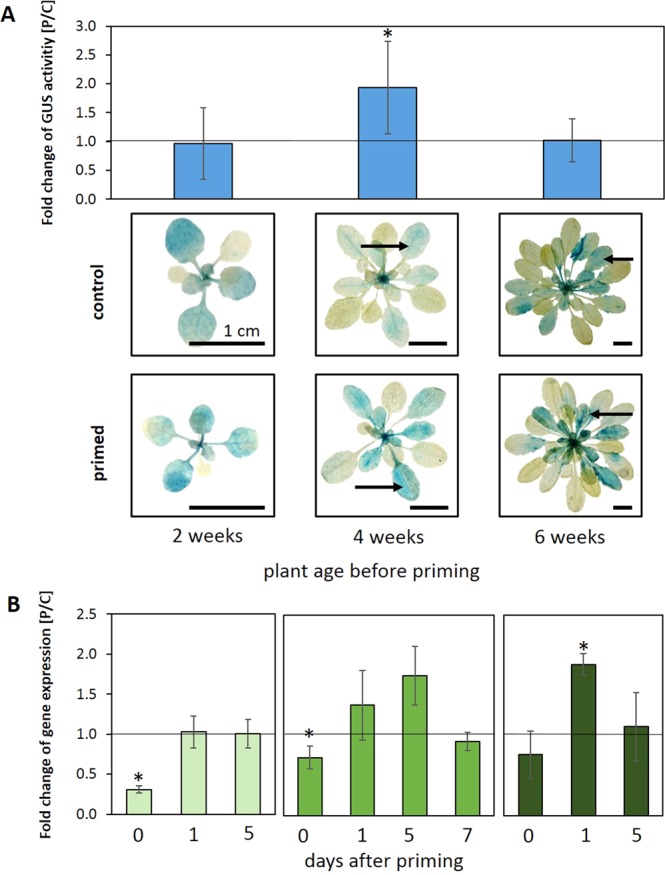
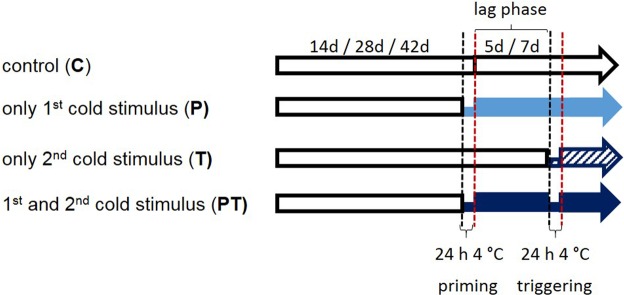
24 h exposure to 4 °C primes Arabidopsis thaliana in the pre-bolting rosette stage for several days against full cold activation of the ROS responsive genes ZAT10 and BAP1 and causes stronger cold-induction of pleiotropically stress-regulated genes. Transient over-expression of thylakoid ascorbate peroxidase (tAPX) at 20 °C mimicked and tAPX transcript silencing antagonized cold-priming of ZAT10 expression. The tAPX effect could not be replaced by over-expression of stromal ascorbate peroxidase (sAPX) demonstrating that priming is specific to regulation of tAPX availability and, consequently, regulated locally at the thylakoid membrane. Arabidopsis acquired cold primability in the early rosette stage between 2 and 4 weeks. During further rosette development, primability was widely maintained in the oldest leaves. Later formed and later maturing leaves were not primable demonstrating that priming is stronger regulated with plant age than with leaf age. In 4-week-old plants, which were strongest primable, the memory was fully erasable and lost seven days after priming. In summary, we conclude that cold-priming of chloroplast-to-nucleus ROS signalling by transient post-stress induction of tAPX transcription is a strategy to modify cell signalling for some time without affecting the alertness for activation of cold acclimation responses.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Thomashow MF. Plant cold acclimation: Freezing tolerance genes and regulatory mechanisms. Annual Review of Plant Physiology and Plant Molecular Biology. 1999;50:571–599. - PubMed
-
- Huner NPA, et al. Photosynthesis, photoinhibition and low-temperature acclimation in cold tolerant plants. Photosynthesis Research. 1993;37:19–39. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
