

Deletion of Seventeen Amino Acids at the C-Terminal End of Aquaporin 0 Causes Distortion Aberration and Cataract in the Lenses of AQP0ΔC/ΔC Mice
- PMID: 30821811
- PMCID: PMC6397018
- DOI: 10.1167/iovs.18-26378
Deletion of Seventeen Amino Acids at the C-Terminal End of Aquaporin 0 Causes Distortion Aberration and Cataract in the Lenses of AQP0ΔC/ΔC Mice
Abstract
Purpose: Investigate the effects of the absence of 17 amino acids at the C-terminal end of Aquaporin 0 (AQP0) on lens transparency, focusing property, and homeostasis.
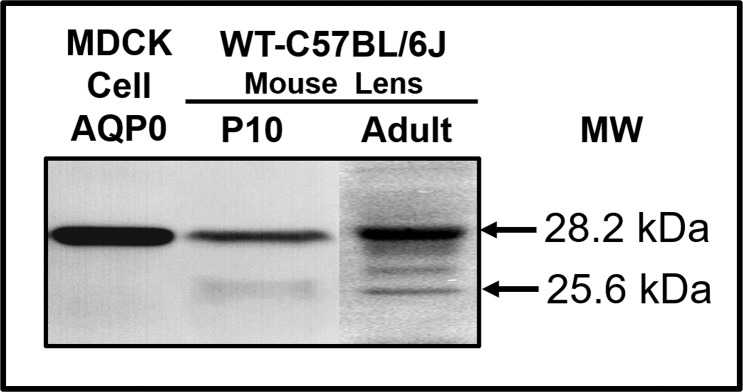
Methods: A knockin (KI) mouse model (AQP0ΔC/ΔC) was developed to express AQP0 only as the end-cleaved form in the lens. For this, AQP0 was genetically engineered as C-terminally end-cleaved with amino acids 1 to 246, instead of the full length 1 to 263 of the wild type (WT). After verifying the KI integration into the genome and its expression, the mouse model was bred for several generations. AQP0 KI homozygous (AQP0ΔC/ΔC) and heterozygous (AQP0+/ΔC) lenses were imaged and analyzed at different developmental stages for transparency. Correspondingly, aberrations in the lens were characterized using the standard metal grid focusing method. Data were compared with age-matched WT, AQP0 knockout (AQP0-/-), and AQP0 heterozygous (AQP0+/-) lenses.
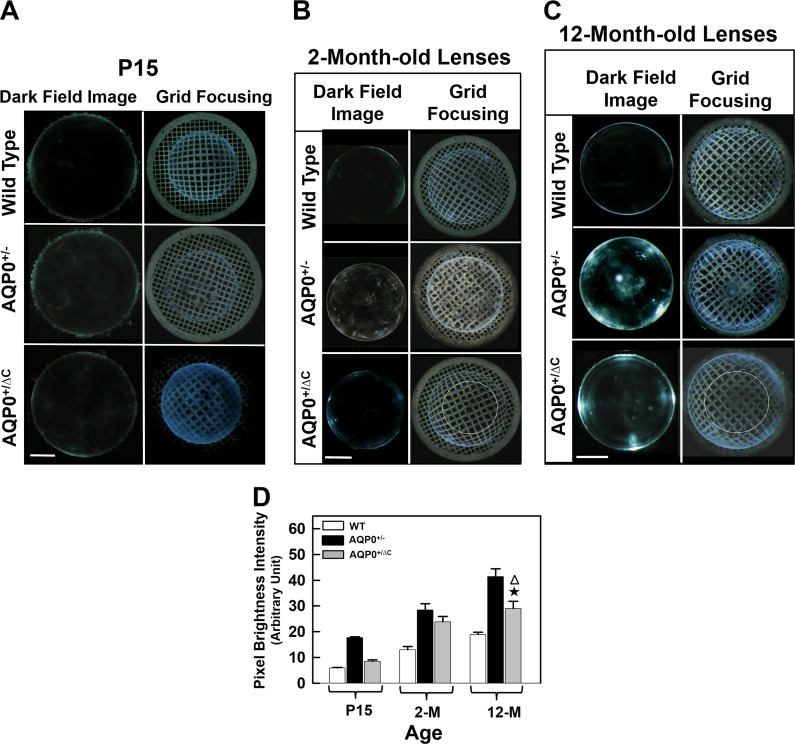
Results: AQP0ΔC/ΔC lenses were transparent throughout the embryonic development and until postnatal day 15 (P15) in contrast to age-matched AQP0-/- lenses, which developed cataract at embryonic stage itself. However, there was distortion aberration in AQP0ΔC/ΔC lens at P5; after P15, cataract began to develop and progressed faster surpassing that of age-matched AQP0-/- lenses. AQP0+/ΔC lenses were transparent even at the age of 1 year in contrast to AQP0+/- lenses; however, there was distortion aberration starting at P15.
Conclusions: A specific distribution profile of intact and end-cleaved AQP0 from the outer cortex to the inner nucleus is required in the lens for establishing refractive index gradient to enable proper focusing without aberrations and for maintaining transparency.
Figures






Similar articles
-
C-Terminal End of Aquaporin 0 Regulates Lens Gap Junction Channel Function.Invest Ophthalmol Vis Sci. 2019 Jun 3;60(7):2525-2531. doi: 10.1167/iovs.19-26787. Invest Ophthalmol Vis Sci. 2019. PMID: 31195409 Free PMC article.
-
A predominant form of C-terminally end-cleaved AQP0 functions as an open water channel and an adhesion protein in AQP0ΔC/ΔC mouse lens.Biochem Biophys Res Commun. 2019 Apr 9;511(3):626-630. doi: 10.1016/j.bbrc.2019.02.098. Epub 2019 Feb 27. Biochem Biophys Res Commun. 2019. PMID: 30826060 Free PMC article.
-
Deletion of beaded filament proteins or the C-terminal end of Aquaporin 0 causes analogous abnormal distortion aberrations in mouse lens.Exp Eye Res. 2021 Aug;209:108645. doi: 10.1016/j.exer.2021.108645. Epub 2021 Jun 1. Exp Eye Res. 2021. PMID: 34087204 Free PMC article.
-
Structural function of MIP/aquaporin 0 in the eye lens; genetic defects lead to congenital inherited cataracts.Handb Exp Pharmacol. 2009;(190):265-97. doi: 10.1007/978-3-540-79885-9_14. Handb Exp Pharmacol. 2009. PMID: 19096783 Review.
-
[Study of the Mechanisms of Maintaining the Transparency of the Lens and Treatment of Its Related Diseases for Making Anti-cataract and/or Anti-presbyopia Drugs].Yakugaku Zasshi. 2020;140(9):1095-1099. doi: 10.1248/yakushi.20-00120. Yakugaku Zasshi. 2020. PMID: 32879241 Review. Japanese.
Cited by
-
Arvcf Dependent Adherens Junction Stability is Required to Prevent Age-Related Cortical Cataracts.Front Cell Dev Biol. 2022 Jul 6;10:840129. doi: 10.3389/fcell.2022.840129. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 35874813 Free PMC article.
-
C-Terminal End of Aquaporin 0 Regulates Lens Gap Junction Channel Function.Invest Ophthalmol Vis Sci. 2019 Jun 3;60(7):2525-2531. doi: 10.1167/iovs.19-26787. Invest Ophthalmol Vis Sci. 2019. PMID: 31195409 Free PMC article.
-
Classification and Gene Structure of Aquaporins.Adv Exp Med Biol. 2023;1398:1-13. doi: 10.1007/978-981-19-7415-1_1. Adv Exp Med Biol. 2023. PMID: 36717483
-
A predominant form of C-terminally end-cleaved AQP0 functions as an open water channel and an adhesion protein in AQP0ΔC/ΔC mouse lens.Biochem Biophys Res Commun. 2019 Apr 9;511(3):626-630. doi: 10.1016/j.bbrc.2019.02.098. Epub 2019 Feb 27. Biochem Biophys Res Commun. 2019. PMID: 30826060 Free PMC article.
-
Lens aquaporins function as peroxiporins to facilitate membrane transport of hydrogen peroxide.Biochem Biophys Res Commun. 2020 Apr 16;524(4):1025-1029. doi: 10.1016/j.bbrc.2020.02.031. Epub 2020 Feb 13. Biochem Biophys Res Commun. 2020. PMID: 32063362 Free PMC article.
References
-
- Pierscionek BK, Regini JW. The gradient index lens of the eye: an opto-biological synchrony. Prog Retin Eye Res. 2012;31:332–349. - PubMed
-
- Kumari SS, Varadaraj K. Intact and N- or C-terminal end truncated AQP0 function as open water channels and cell-to-cell adhesion proteins: end truncation could be a prelude for adjusting the refractive index of the lens to prevent spherical aberration. Biochim Biophys Acta. 2014;1840:2862–2877. - PMC - PubMed
-
- Hecht E. Optics 2nd ed. Boston: Addison Wesley;; 1987.
-
- Garner LF, Ooi CS, Smith G. Refractive index of the crystalline lens in young and aged eyes. Clin Exp Optom. 1998;81:145–150. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
