

Caffeine inhibits hypoxia-induced nuclear accumulation in HIF-1α and promotes neonatal neuronal survival
- PMID: 30822423
- PMCID: PMC6935249
- DOI: 10.1016/j.expneurol.2019.01.014
Caffeine inhibits hypoxia-induced nuclear accumulation in HIF-1α and promotes neonatal neuronal survival
Abstract
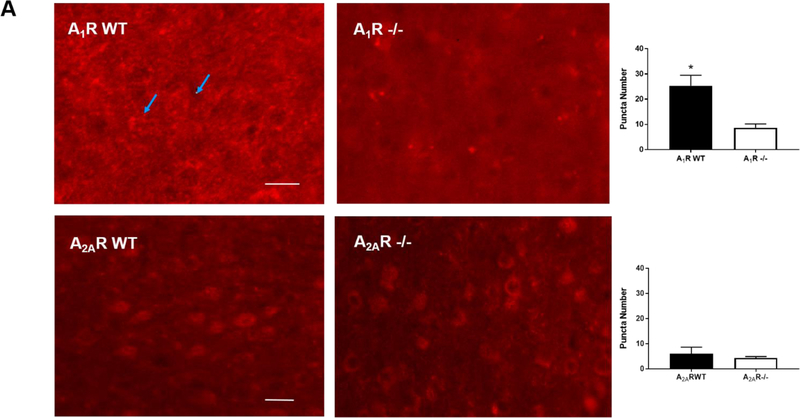
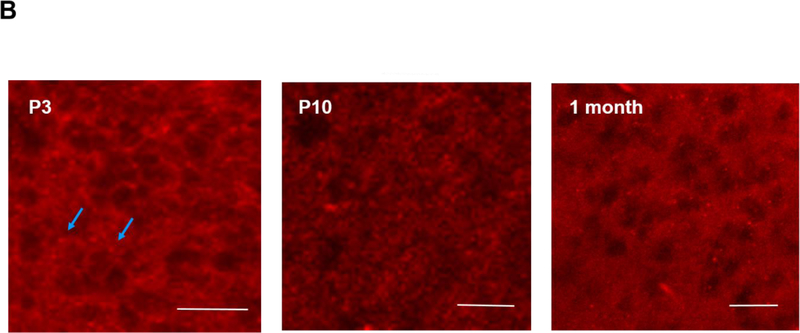
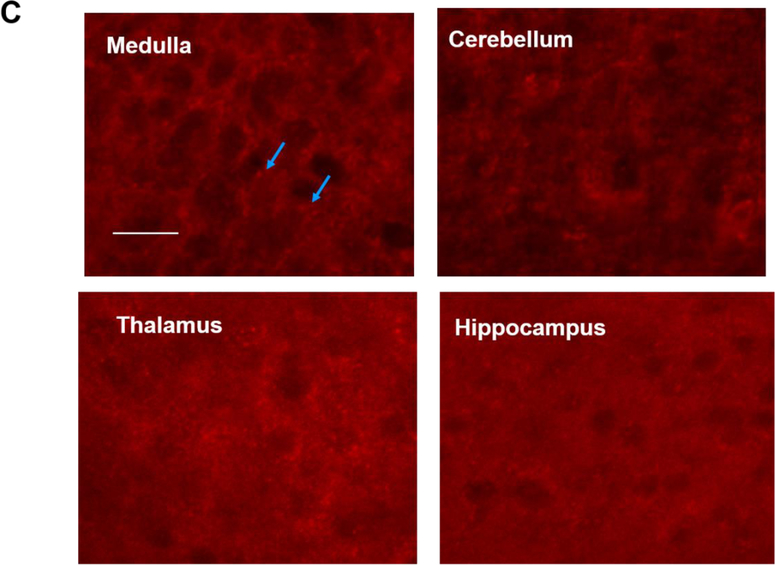
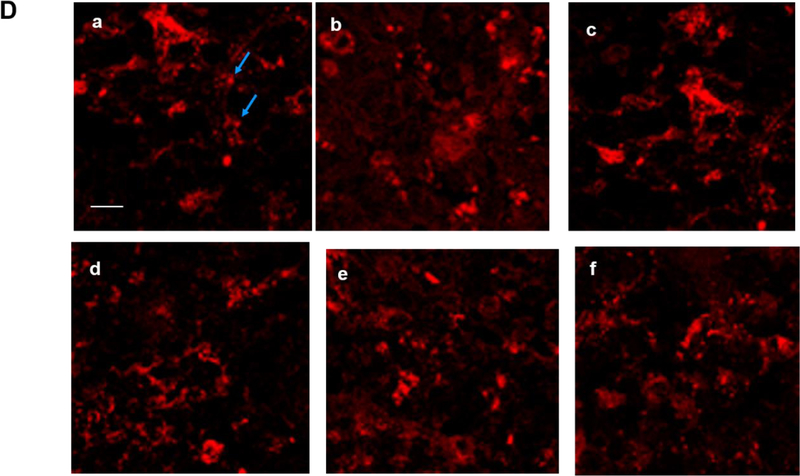
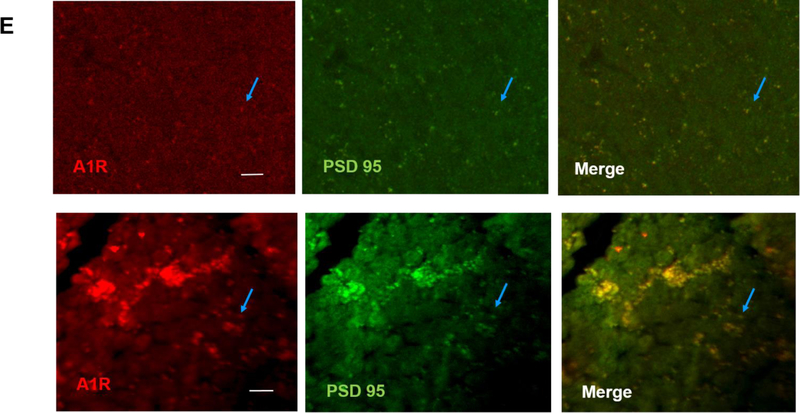
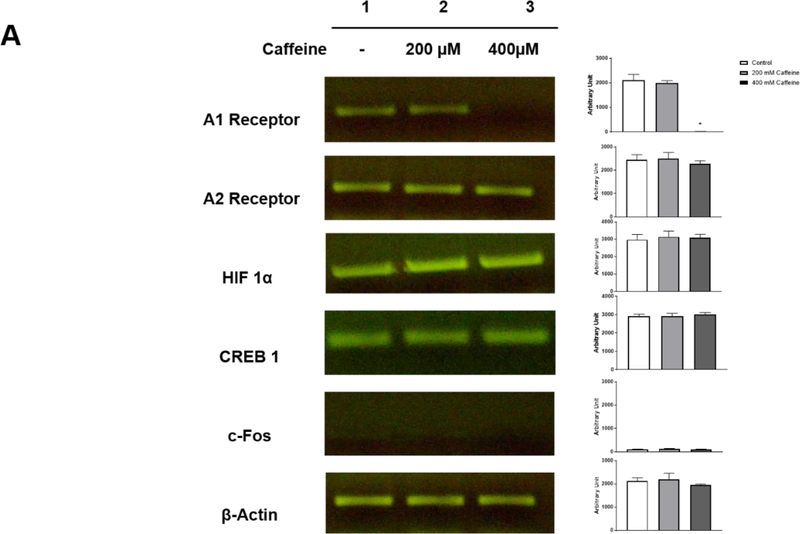
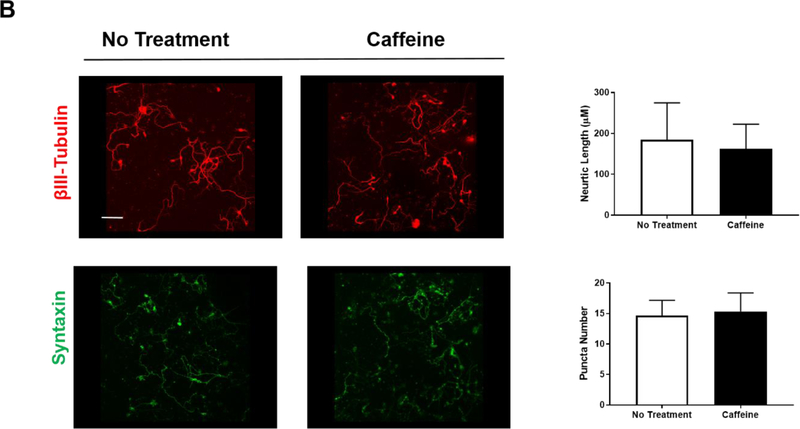
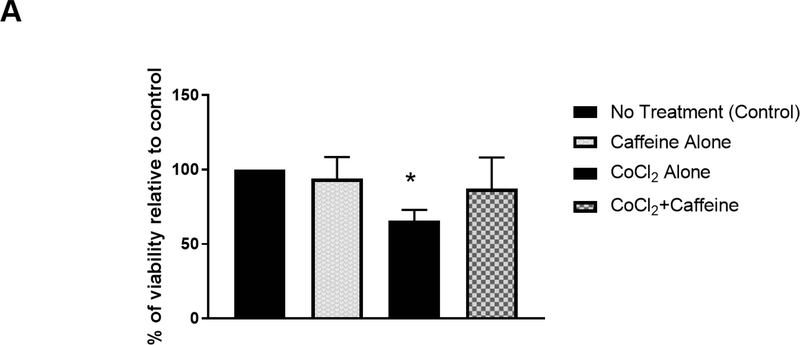
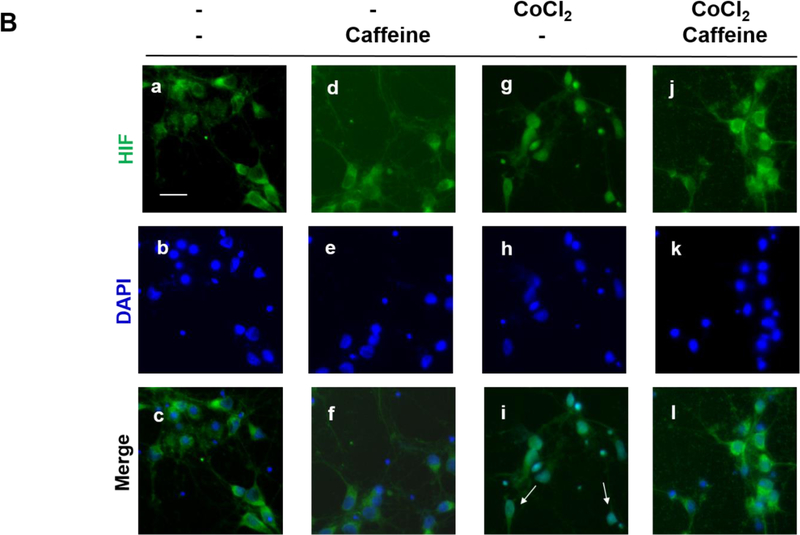
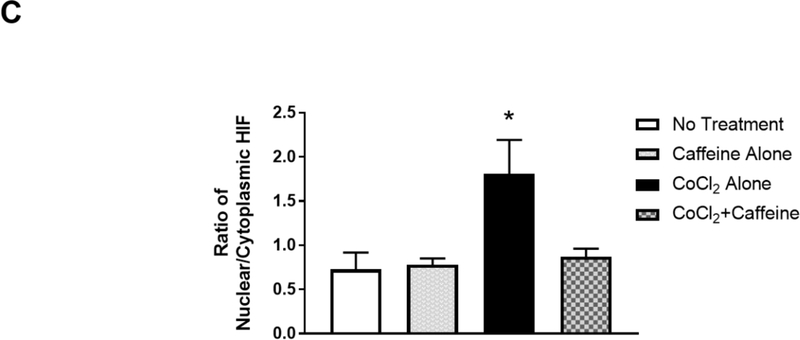
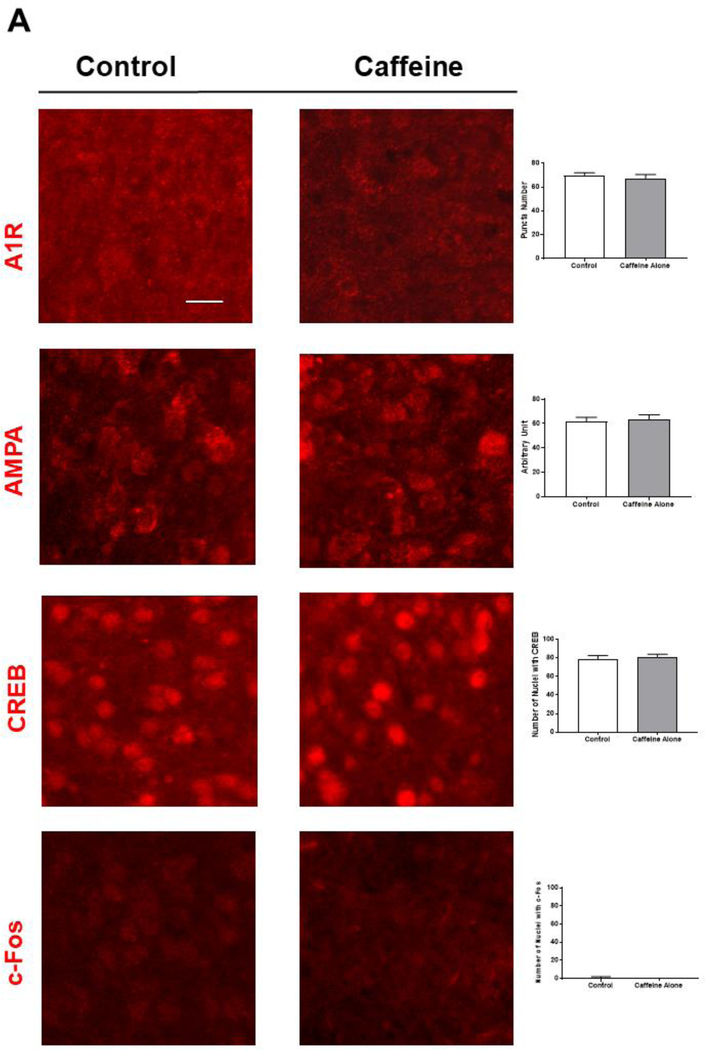
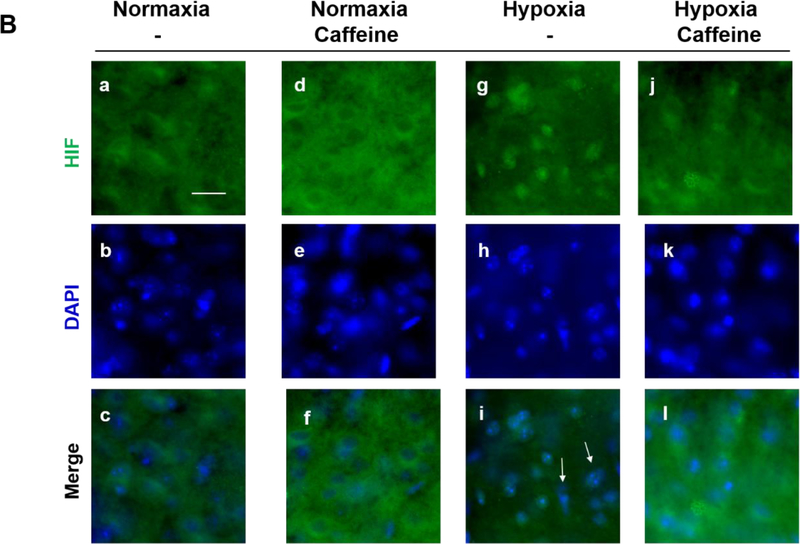
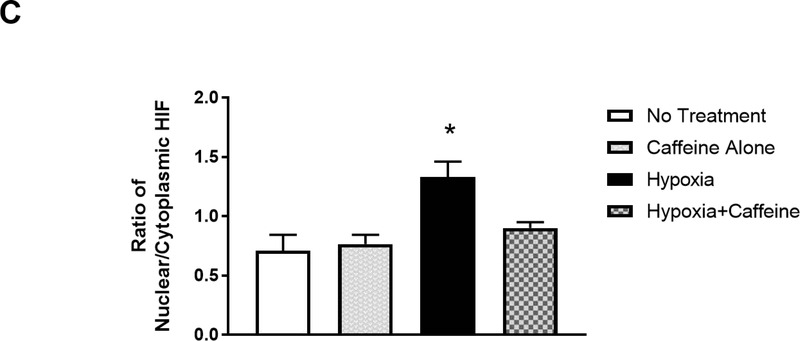
Apnea of prematurity (AOP) defined as cessation of breathing for 15-20 s, is commonly seen in preterm infants. Caffeine is widely used to treat AOP due to its safety and effectiveness. Caffeine releases respiratory arrest by competing with adenosine for binding to adenosine A1 and A2A receptors (A1R and A2AR). Long before its use in treating AOP, caffeine has been used as a psychostimulant in adult brains. However, the effect of caffeine on developing brains remains unclear. We found that A1R proteins for caffeine binding were expressed in the brains of neonatal rodents and preterm infants (26-27 weeks). Neonatal A1R proteins colocalized with PSD-95, suggesting its synaptic localization. In contrast, our finding on A2R expression in neonatal neurons was restricted to the mRNA level as detected by single cell RT/PCR due to the lack of specific A2AR antibody. Furthermore, caffeine (200 μM) at a dose twice higher than the clinically relevant dose (36-130 μM) had minor or no effects on several basic neuronal functions, such as neurite outgrowth, synapse formation, expression of A1R and transcription of CREB-1 and c-Fos, further supporting the safety of caffeine for clinical use. We found that treatment with CoCl2 (125 μM), a hypoxia mimetic agent, for 24 h triggered neuronal death and nuclear accumulation of HIF-1α in primary neuronal cultures. Subsequent treatment with caffeine at a concentration of 100 μM alleviated CoCl2-induced cell death and prevented nuclear accumulation of HIF-1α. Consistently, caffeine treatment in early postnatal life of neonatal mice (P4-P7) also prevented subsequent hypoxia-induced nuclear increase of HIF-1α. Together, our data support the utility of caffeine in alleviating hypoxia-induced damages in developing neurons.
Copyright © 2019 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of Interest
All authors have read the journal’s policy on disclosure of potential conflicts of interest and have none to declare. This study was supported by two non-profit foundations. The funders played no role in the design and conclusion of this study.
Figures
References
-
- Aden U, 2011. Methylxanthines during pregnancy and early postnatal life. Handb Exp Pharmacol, 373–389. - PubMed
-
- Aden U, Herlenius E, Tang LQ, Fredholm BB, 2000. Maternal caffeine intake has minor effects on adenosine receptor ontogeny in the rat brain. Pediatr Res 48, 177–183. - PubMed
-
- Aden U, Leverin AL, Hagberg H, Fredholm BB, 2001. Adenosine A(1) receptor agonism in the immature rat brain and heart. Eur J Pharmacol 426, 185–192. - PubMed
-
- Back SA, Craig A, Luo NL, Ren J, Akundi RS, Ribeiro I, Rivkees SA, 2006. Protective effects of caffeine on chronic hypoxia-induced perinatal white matter injury. Ann Neurol 60, 696–705. - PubMed
-
- Bairam A, Boutroy MJ, Badonnel Y, Vert P, 1987. Theophylline versus caffeine: comparative effects in treatment of idiopathic apnea in the preterm infant. J Pediatr 110, 636–639. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
