

Inhibition of ricin A-chain (RTA) catalytic activity by a viral genome-linked protein (VPg)
- PMID: 30822539
- PMCID: PMC7219317
- DOI: 10.1016/j.bbapap.2019.02.002
Inhibition of ricin A-chain (RTA) catalytic activity by a viral genome-linked protein (VPg)
Abstract
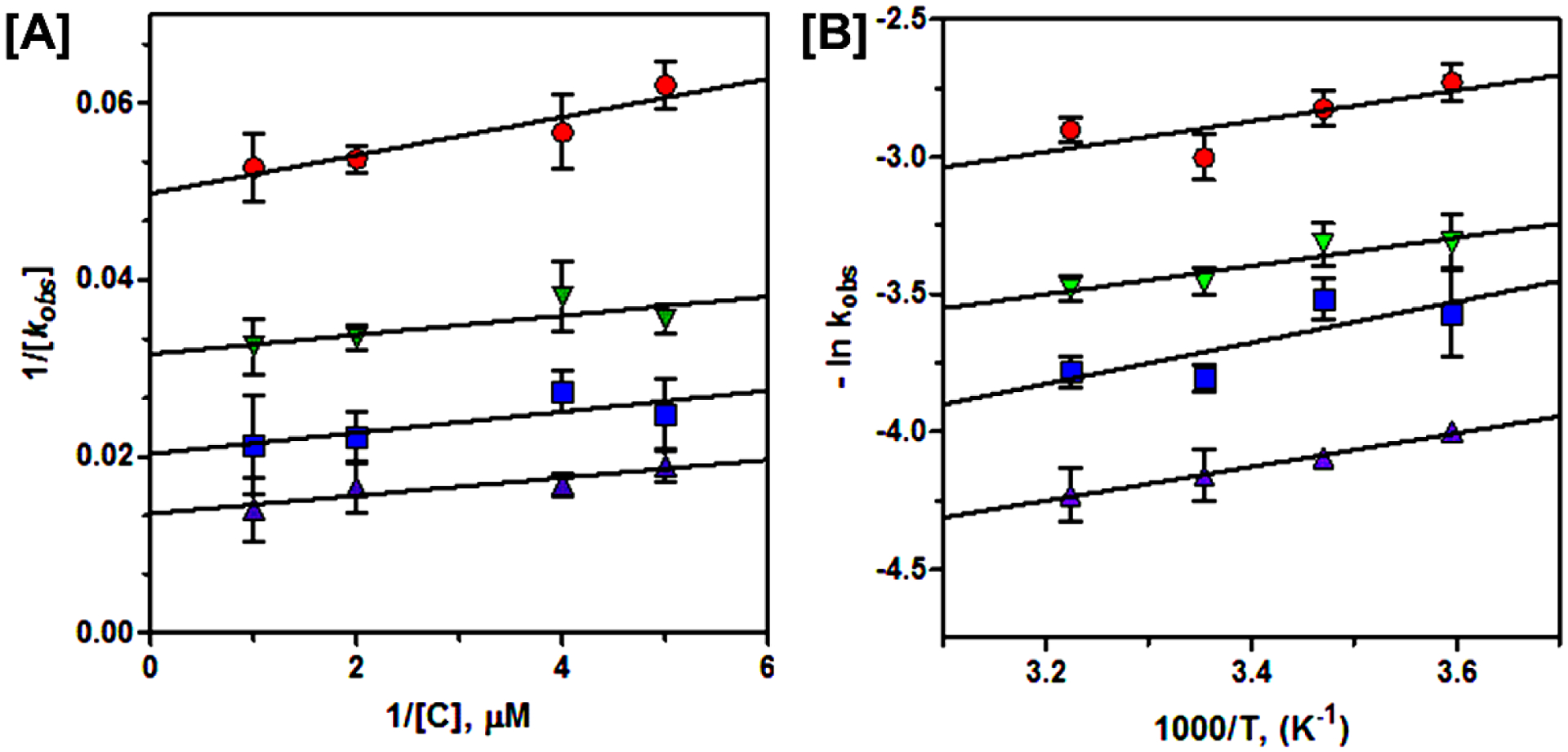
Ricin is a plant derived protein toxin produced by the castor bean plant (Ricinus communis). The Centers for Disease Control (CDC) classifies ricin as a Category B biological agent. Currently, there is neither an effective vaccine that can be used to protect against ricin exposure nor a therapeutic to reverse the effects once exposed. Here we quantitatively characterize interactions between catalytic ricin A-chain (RTA) and a viral genome-linked protein (VPg) from turnip mosaic virus (TuMV). VPg and its N-terminal truncated variant, VPg1-110, bind to RTA and abolish ricin's catalytic depurination of 28S rRNA in vitro and in a cell-free rabbit reticulocyte translational system. RTA and VPg bind in a 1 to 1 stoichiometric ratio, and their binding affinity increases ten-fold as temperature elevates (5 °C to 37 °C). RTA-VPg binary complex formation is enthalpically driven and favored by entropy, resulting in an overall favorable energy, ΔG = -136.8 kJ/mol. Molecular modeling supports our experimental observations and predicts a major contribution of electrostatic interactions, suggesting an allosteric mechanism of downregulation of RTA activity through conformational changes in RTA structure, and/or disruption of binding with the ribosomal stalk. Fluorescence anisotropy studies show that heat affects the rate constant and the activation energy for the RTA-VPg complex, Ea = -62.1 kJ/mol. The thermodynamic and kinetic findings presented here are an initial lead study with promising results and provides a rational approach for synthesis of therapeutic peptides that successfully eliminate toxicity of ricin, and other cytotoxic RIPs.
Keywords: Depurination; Fluorescence; Genome-linked viral protein (VPg); Protein-protein interactions; Ribosome inactivating protein (RIP); Ricin A-chain (RTA).
Copyright © 2019. Published by Elsevier B.V.
Figures







References
-
- Barbieri L, Battelli MG, and Stirpe F (1993) Ribosome-inactivating proteins from plants. Biochim. Biophys. Acta 1154, 237–282. - PubMed
-
- Wang P and Tumer NE (2000) Virus resistance mediated by ribosome inactivating proteins. Adv. Virus Res 55, 325–355. - PubMed
-
- Jimenez A and Vazquez D (1985) Plant and fungal protein and glycoprotein toxins inhibiting eukaryote protein synthesis. Annu. Rev. Microbiol 39, 649–672. - PubMed
-
- Stirpe F (2004) Ribosome-inactivating proteins. Toxicon 44, 371–383. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
