

Hemizygosity Enables a Mutational Transition Governing Fungal Virulence and Commensalism
- PMID: 30824263
- PMCID: PMC6624852
- DOI: 10.1016/j.chom.2019.01.005
Hemizygosity Enables a Mutational Transition Governing Fungal Virulence and Commensalism
Abstract
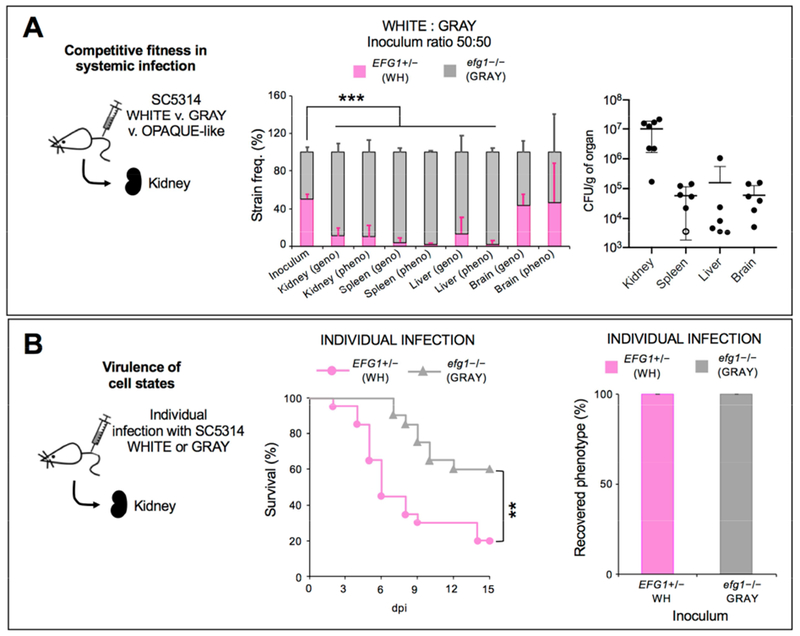
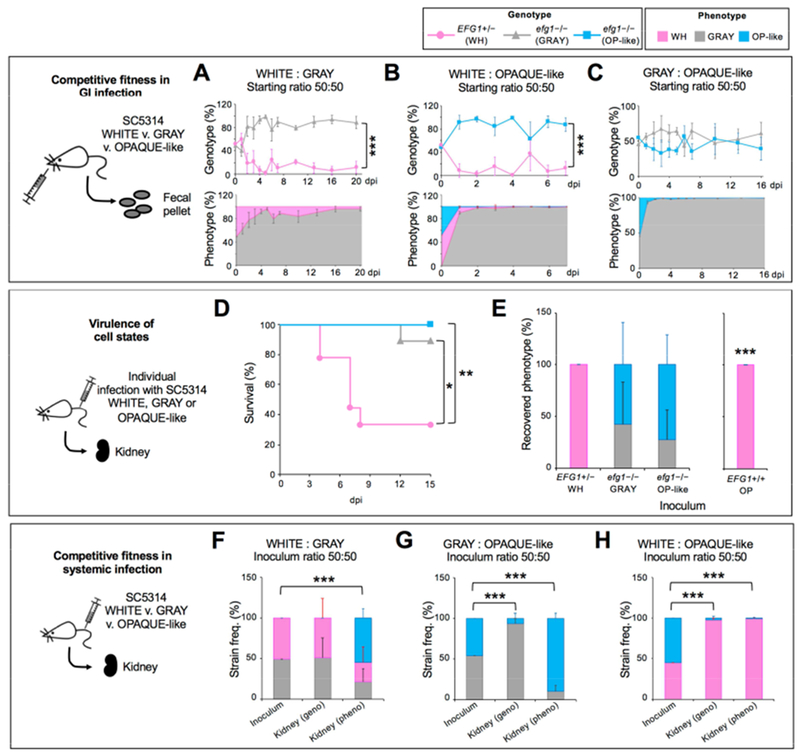
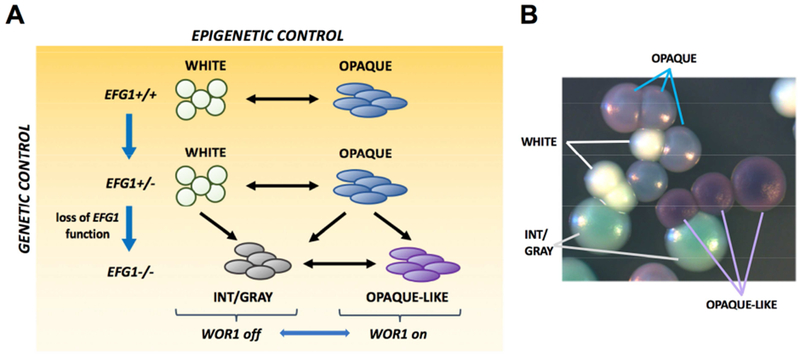
Candida albicans is a commensal fungus of human gastrointestinal and reproductive tracts, but also causes life-threatening systemic infections. The balance between colonization and pathogenesis is associated with phenotypic plasticity, with alternative cell states producing different outcomes in a mammalian host. Here, we reveal that gene dosage of a master transcription factor regulates cell differentiation in diploid C. albicans cells, as EFG1 hemizygous cells undergo a phenotypic transition inaccessible to "wild-type" cells with two functional EFG1 alleles. Notably, clinical isolates are often EFG1 hemizygous and thus licensed to undergo this transition. Phenotypic change corresponds to high-frequency loss of the functional EFG1 allele via de novo mutation or gene conversion events. This phenomenon also occurs during passaging in the gastrointestinal tract with the resulting cell type being hypercompetitive for commensal and systemic infections. A "two-hit" genetic model therefore underlies a key phenotypic transition in C. albicans that enables adaptation to host niches.
Keywords: Candida albicans; gastrointestinal tract; pathogenesis; phase variation; phenotypic switch; population genetics; transcription factor.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interests.
Figures
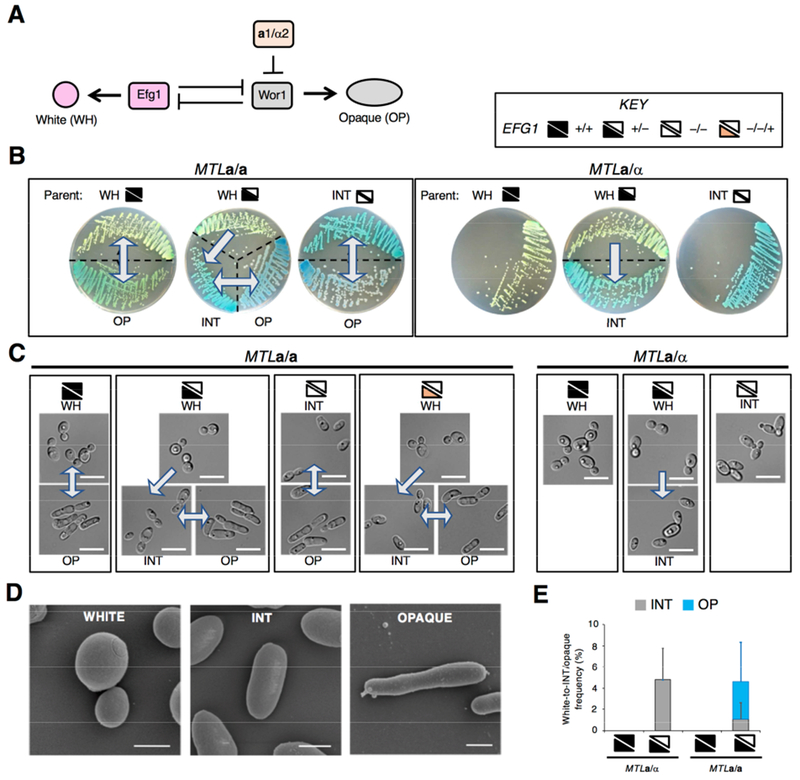
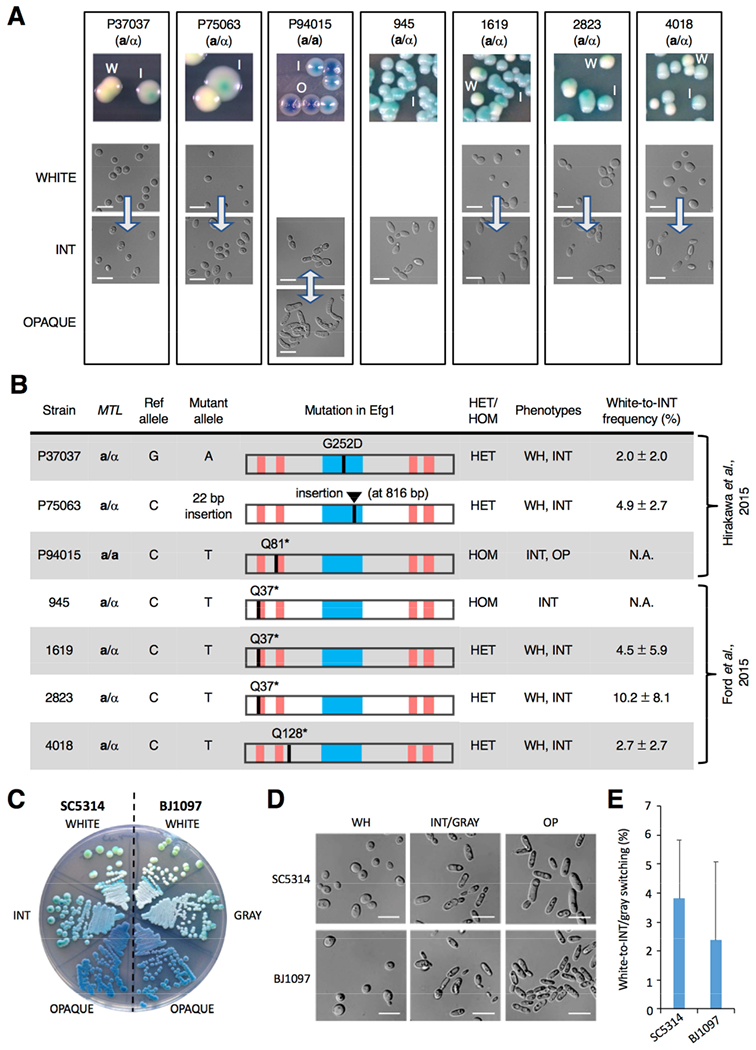
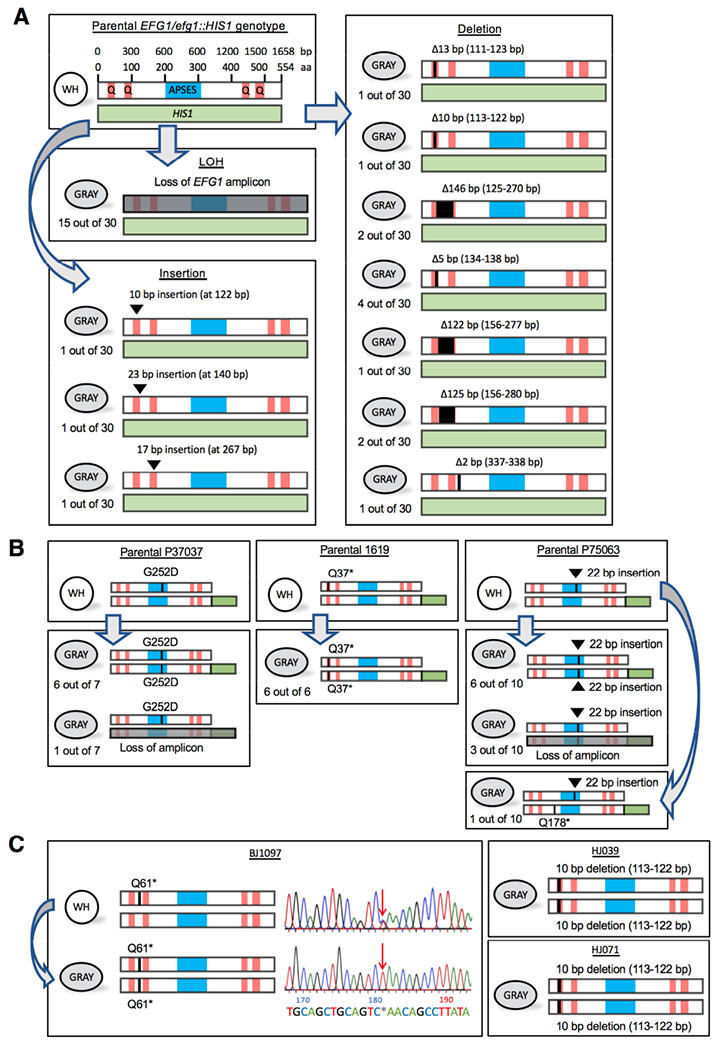
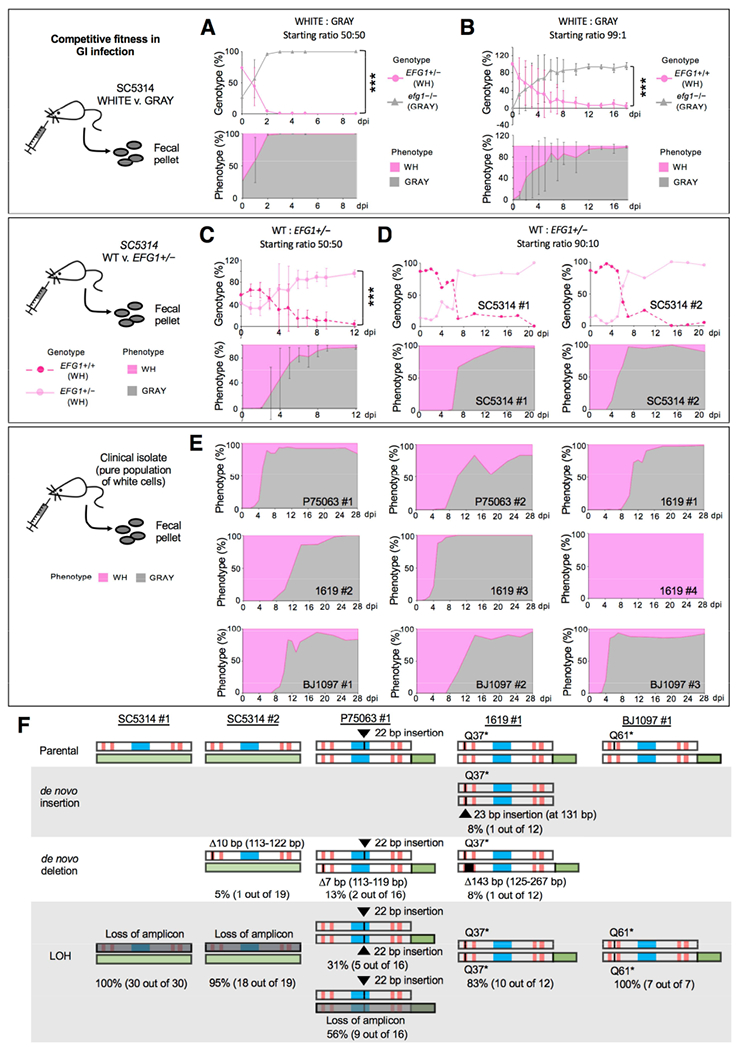
References
-
- Ackermann M (2015). A functional perspective on phenotypic heterogeneity in microorganisms. Nat. Rev. Microbiol 13, 497–508. - PubMed
-
- Bendel CM, Hess DJ, Garni RM, Henry-Stanley M, and Wells CL (2003). Comparative virulence of Candida albicans yeast and filamentous forms in orally and intravenously inoculated mice. Critical Care Med. 31, 501–507. - PubMed
-
- Bennett RJ, and Johnson AD (2006). The role of nutrient regulation and the Gpa2 protein in the mating pheromone response of C. albicans. Mol. Microbiol 62, 100–119. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
