

RNA Binding Antagonizes Neurotoxic Phase Transitions of TDP-43
- PMID: 30826182
- PMCID: PMC6472983
- DOI: 10.1016/j.neuron.2019.01.048
RNA Binding Antagonizes Neurotoxic Phase Transitions of TDP-43
Abstract
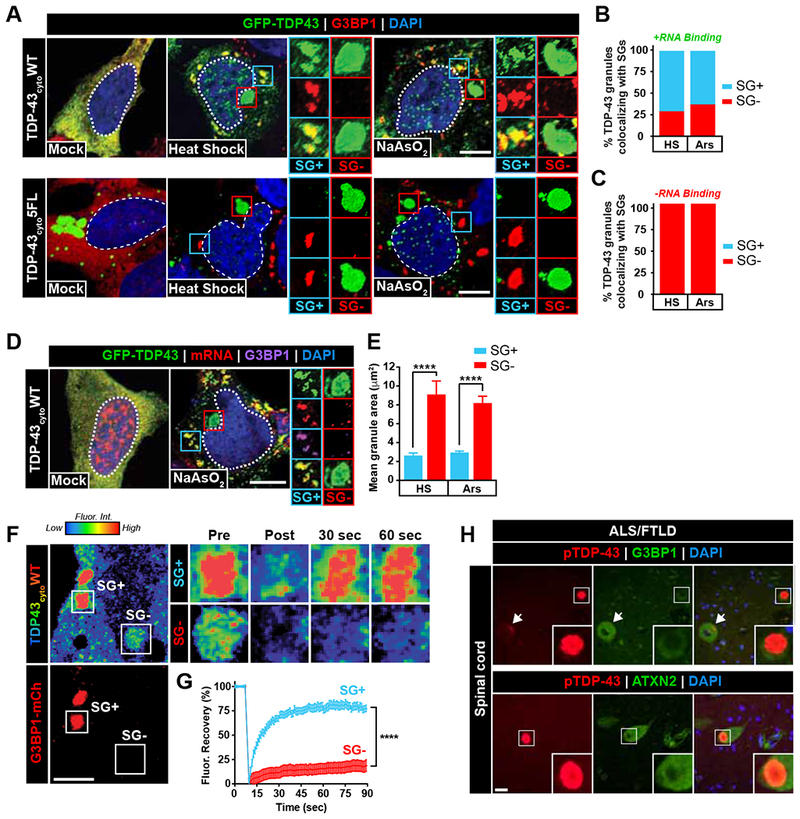
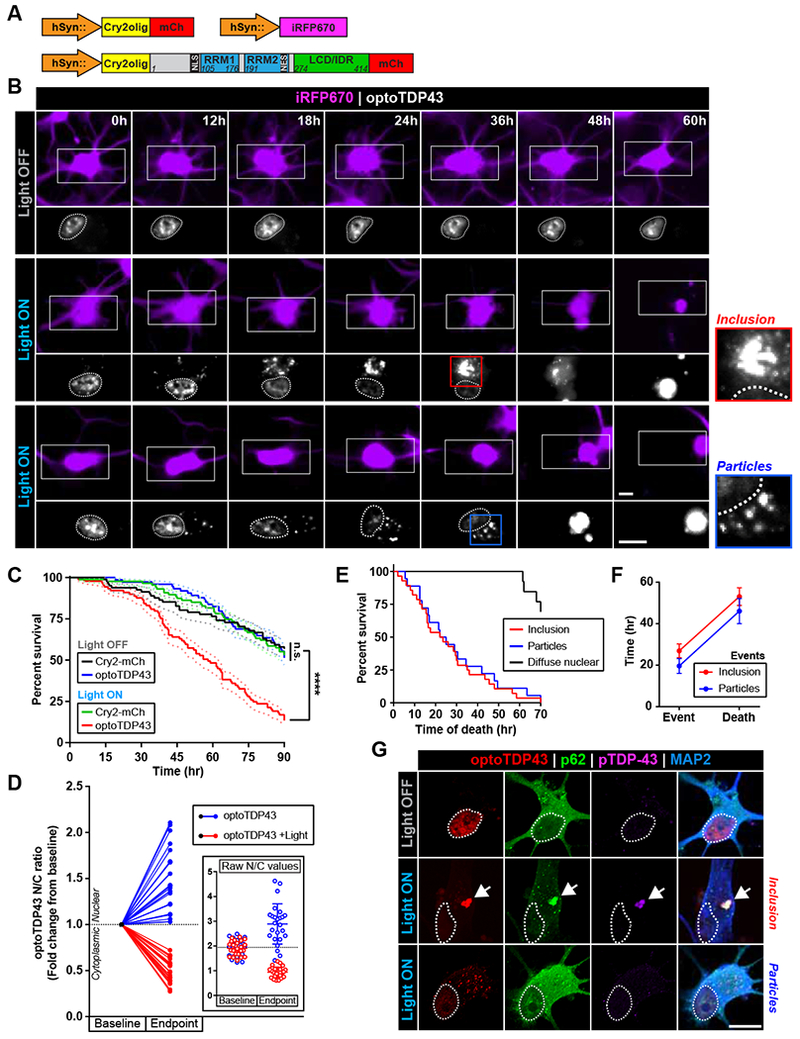
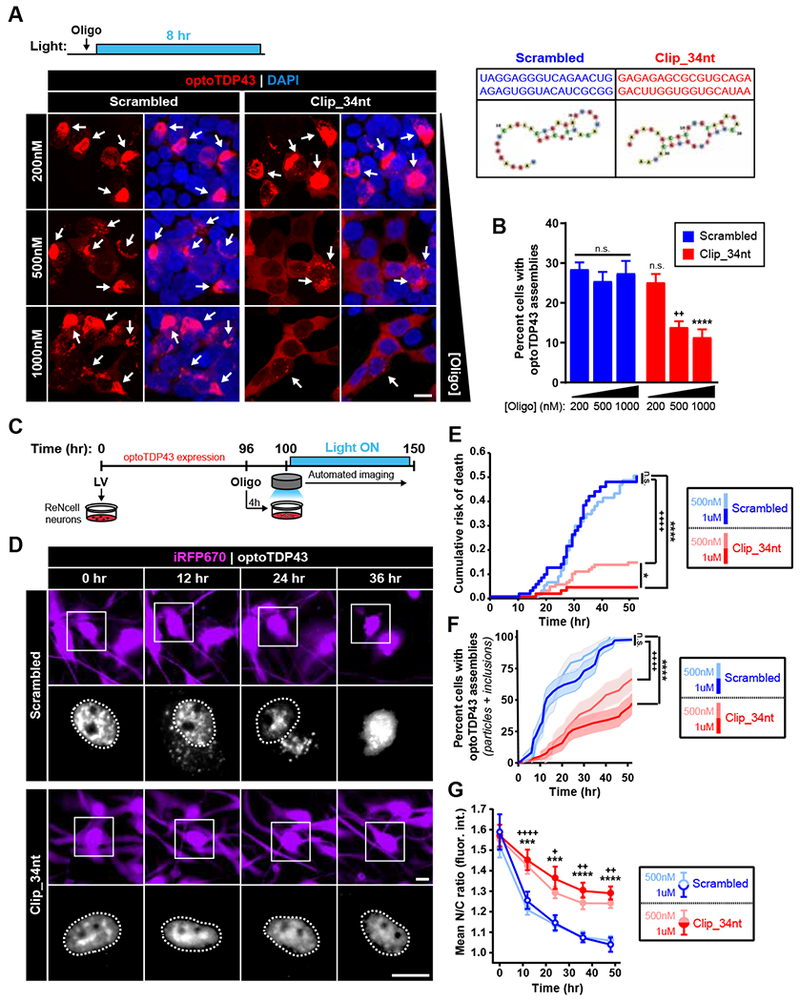
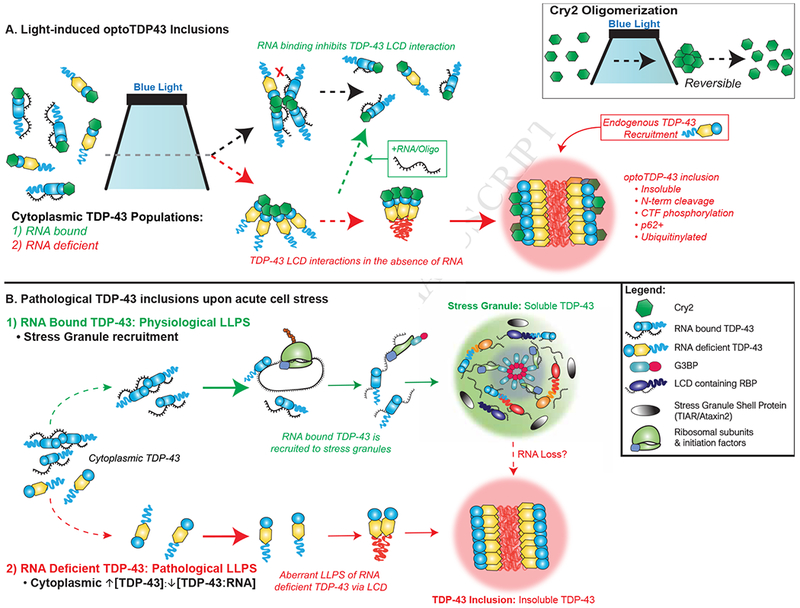
TDP-43 proteinopathy is a pathological hallmark of amyotrophic lateral sclerosis and frontotemporal dementia where cytoplasmic TDP-43 inclusions are observed within degenerating regions of patient postmortem tissue. The mechanism by which TDP-43 aggregates has remained elusive due to technological limitations, which prevent the analysis of specific TDP-43 interactions in live cells. We present an optogenetic approach to reliably induce TDP-43 proteinopathy under spatiotemporal control. We show that the formation of pathologically relevant inclusions is driven by aberrant interactions between low-complexity domains of TDP-43 that are antagonized by RNA binding. Although stress granules are hypothesized to be a conduit for seeding TDP-43 proteinopathy, we demonstrate pathological inclusions outside these RNA-rich structures. Furthermore, we show that aberrant phase transitions of cytoplasmic TDP-43 are neurotoxic and that treatment with oligonucleotides composed of TDP-43 target sequences prevent inclusions and rescue neurotoxicity. Collectively, these studies provide insight into the mechanisms that underlie TDP-43 proteinopathy and present a potential avenue for therapeutic intervention.
Keywords: ALS; FTD; LLPS; RBP; RNA binding protein; TDP-43; bait oligonucleotide; liquid-liquid phase separation; neurodegeneration; optoTDP43; proteinopathy; stress granule.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
Competing Interests
No competing financial interests to declare. Dr. Christopher Donnelly and Jacob Mann are inventors of intellectual property described in this manuscript and submitted as provisional patents by the University of Pittsburgh.
Figures
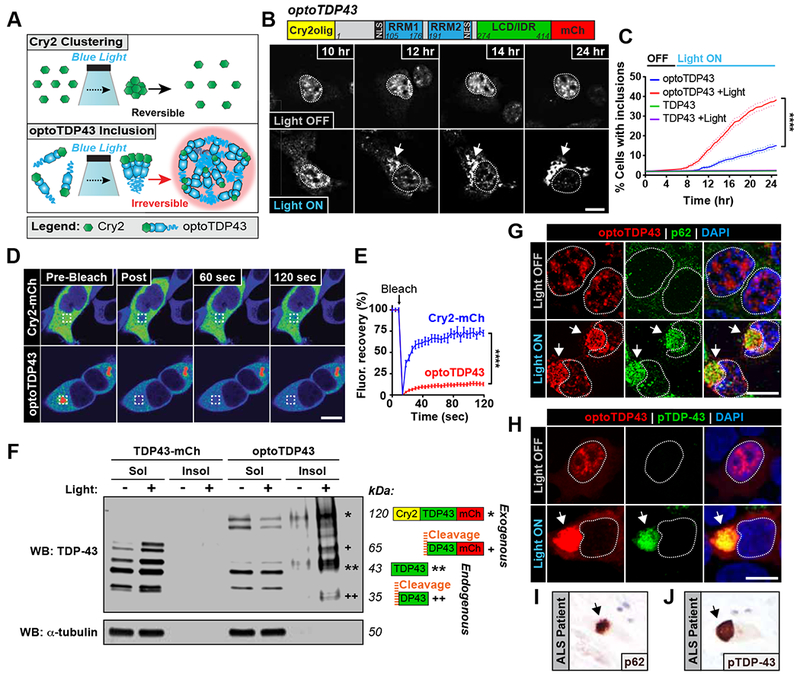
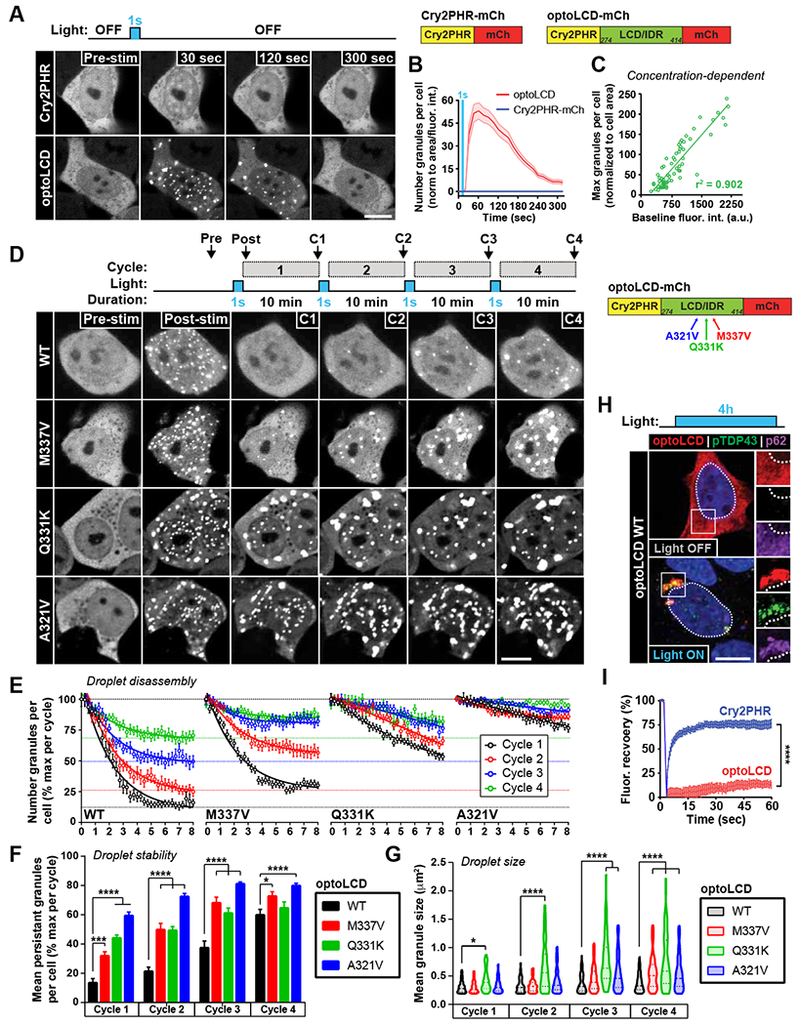
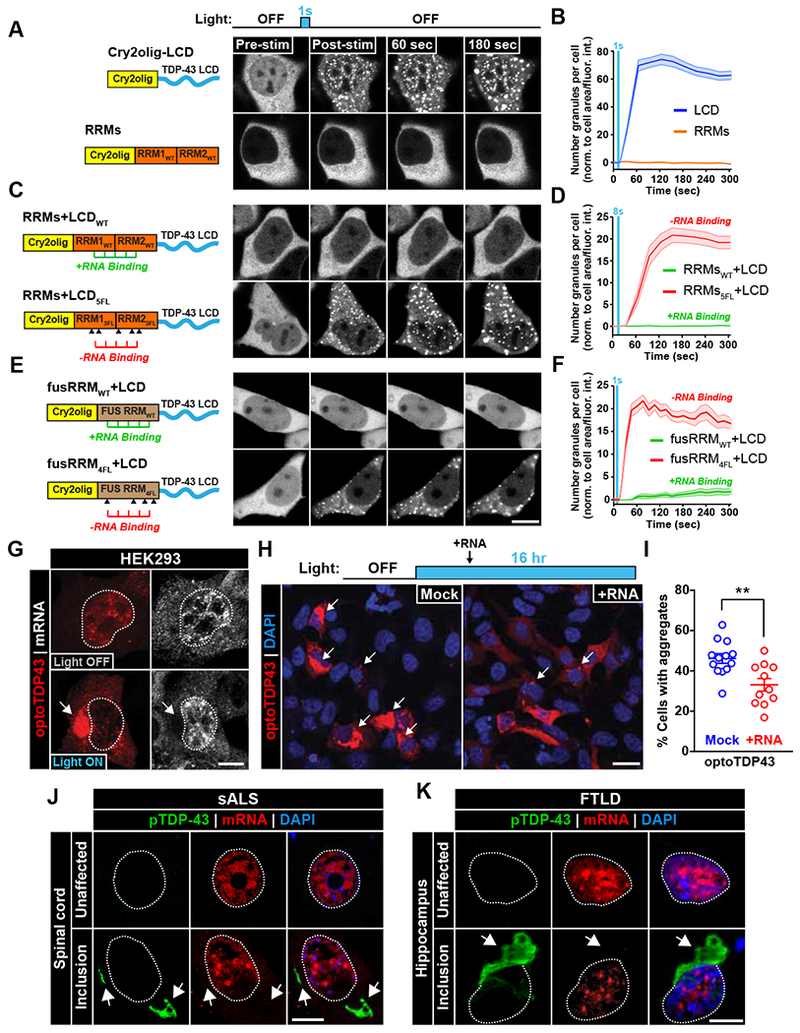
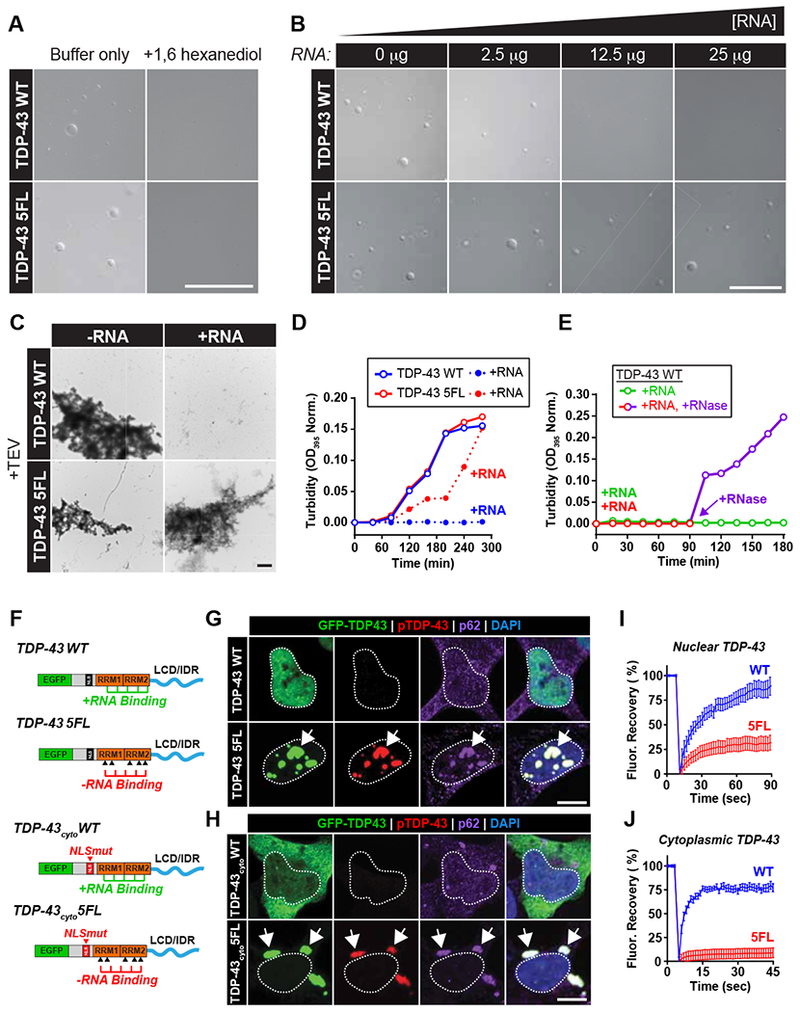
Comment in
-
The Evolution of Phase-Separated TDP-43 in Stress.Neuron. 2019 Apr 17;102(2):265-267. doi: 10.1016/j.neuron.2019.03.041. Neuron. 2019. PMID: 30998893 Free PMC article.
References
-
- Al-Sarraj S, King A, Troakes C, Smith B, Maekawa S, Bodi I, Rogelj B, Al-Chalabi A, Hortobágyi T, and Shaw CE (2011). p62 positive, TDP-43 negative, neuronal cytoplasmic and intranuclear inclusions in the cerebellum and hippocampus define the pathology of C9orf72-linked FTLD and MND/ALS. Acta Neuropathol 122, 691–702. - PubMed
-
- Anderson P, and Kedersha N (2008). Stress granules: the Tao of RNA triage. Trends Biochem. Sci 33, 141–150. - PubMed
-
- Arnold ES, Ling S-C, Huelga SC, Lagier-Tourenne C, Polymenidou M, Ditsworth D, Kordasiewicz HB, McAlonis-Downes M, Platoshyn O, Parone PA, et al. (2013). ALS-linked TDP-43 mutations produce aberrant RNA splicing and adult-onset motor neuron disease without aggregation or loss of nuclear TDP-43. Proc. Natl. Acad. Sci. USA 110, E736–45. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
