

Megabase Length Hypermutation Accompanies Human Structural Variation at 17p11.2
- PMID: 30827684
- PMCID: PMC6438178
- DOI: 10.1016/j.cell.2019.01.045
Megabase Length Hypermutation Accompanies Human Structural Variation at 17p11.2
Abstract
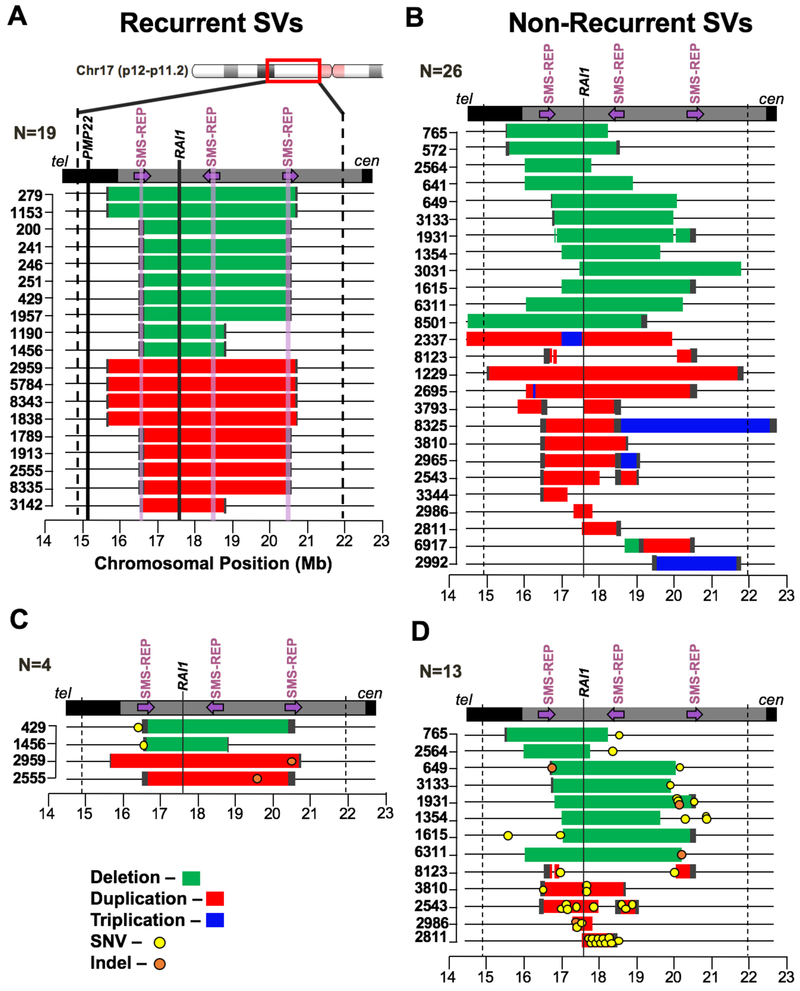
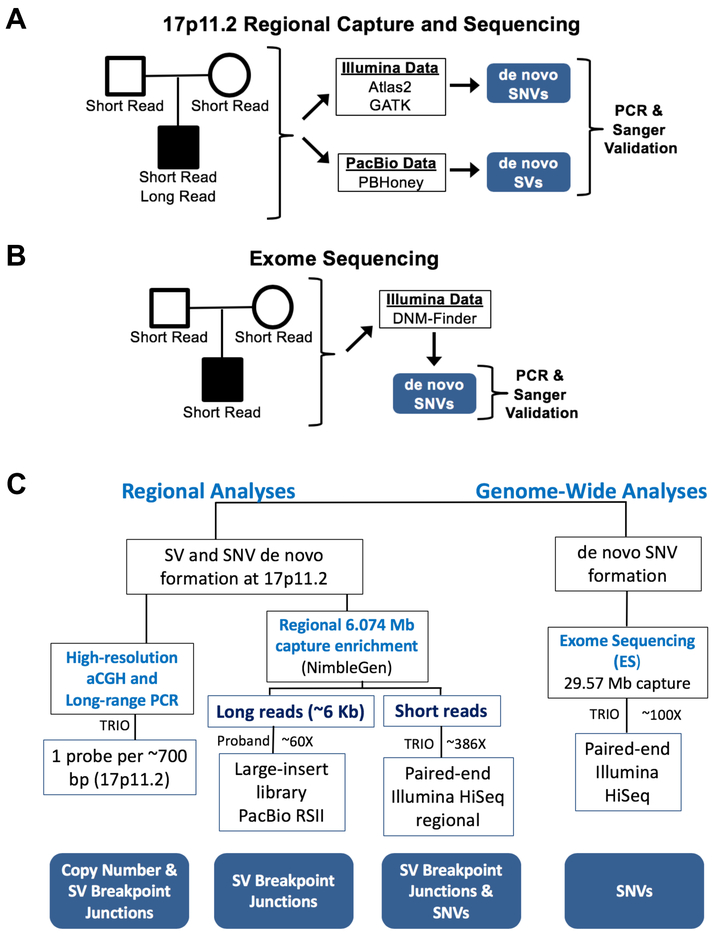
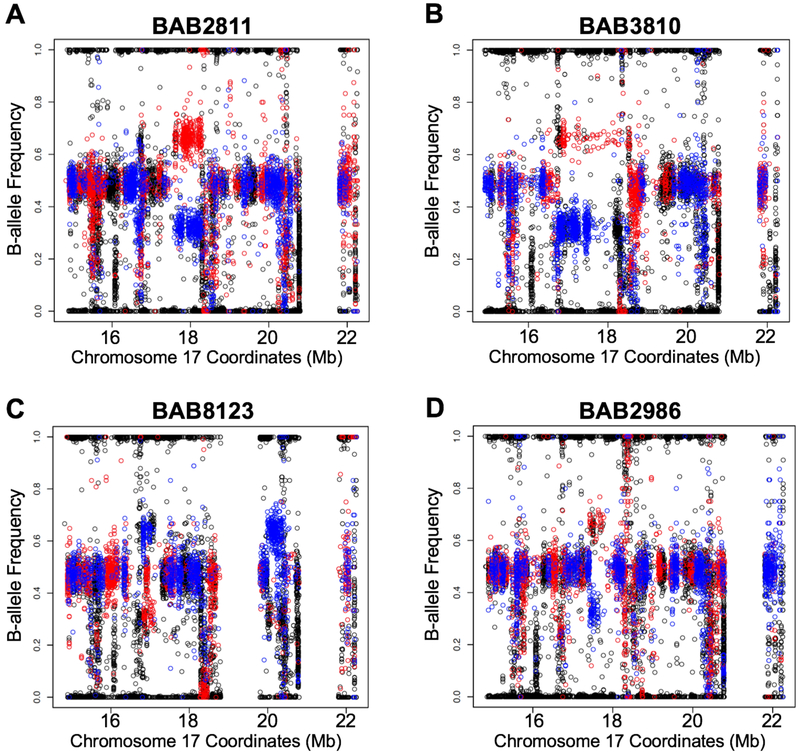
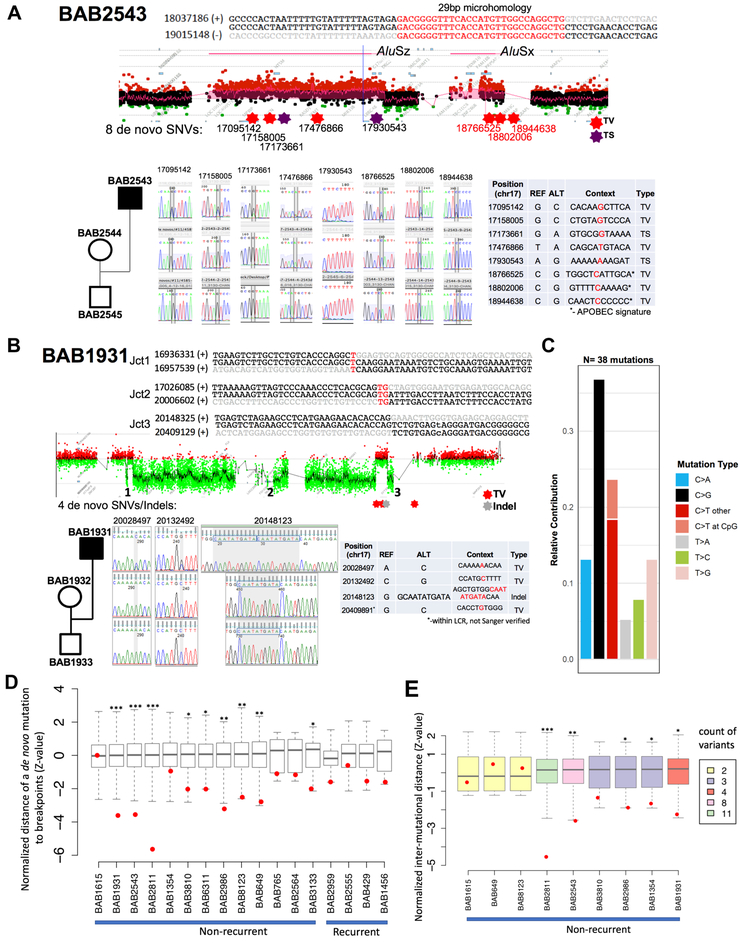
DNA rearrangements resulting in human genome structural variants (SVs) are caused by diverse mutational mechanisms. We used long- and short-read sequencing technologies to investigate end products of de novo chromosome 17p11.2 rearrangements and query the molecular mechanisms underlying both recurrent and non-recurrent events. Evidence for an increased rate of clustered single-nucleotide variant (SNV) mutation in cis with non-recurrent rearrangements was found. Indel and SNV formation are associated with both copy-number gains and losses of 17p11.2, occur up to ∼1 Mb away from the breakpoint junctions, and favor C > G transversion substitutions; results suggest that single-stranded DNA is formed during the genesis of the SV and provide compelling support for a microhomology-mediated break-induced replication (MMBIR) mechanism for SV formation. Our data show an additional mutational burden of MMBIR consisting of hypermutation confined to the locus and manifesting as SNVs and indels predominantly within genes.
Keywords: CNVs; DNA repair; complex rearrangements; genomic characterization; genomic disorders; long-read sequencing; phasing.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
Baylor College of Medicine (BCM) and Miraca Holdings have formed a joint venture with shared ownership and governance of the Baylor Genetics (BG), which performs clinical microarray analysis and clinical exome sequencing. C.A.S. is an employee of BCM and derives support through a professional services agreement with the BG. J.R.L. serves on the Scientific Advisory Board of the BG. J.R.L. has stock ownership in 23andMe, is a paid consultant for Regeneron Pharmaceuticals, has stock options in Lasergen, and is a co-inventor on multiple United States and European patents related to molecular diagnostics for inherited neuropathies, eye diseases, and bacterial genomic fingerprinting.
Figures
References
Publication types
MeSH terms
Supplementary concepts
Grants and funding
- P30 CA016672/CA/NCI NIH HHS/United States
- R35 GM133600/GM/NIGMS NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- R35 NS105078/NS/NINDS NIH HHS/United States
- T32 NS043124/NS/NINDS NIH HHS/United States
- K99 GM120453/GM/NIGMS NIH HHS/United States
- R01 NS058529/NS/NINDS NIH HHS/United States
- R00 GM120453/GM/NIGMS NIH HHS/United States
- UM1 HG006542/HG/NHGRI NIH HHS/United States
- U54 HG003273/HG/NHGRI NIH HHS/United States
- R01 GM080600/GM/NIGMS NIH HHS/United States
- R01 GM106373/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
