

Emergence of a Chimeric Globin Pseudogene and Increased Hemoglobin Oxygen Affinity Underlie the Evolution of Aquatic Specializations in Sirenia
- PMID: 30828717
- PMCID: PMC6526914
- DOI: 10.1093/molbev/msz044
Emergence of a Chimeric Globin Pseudogene and Increased Hemoglobin Oxygen Affinity Underlie the Evolution of Aquatic Specializations in Sirenia
Abstract
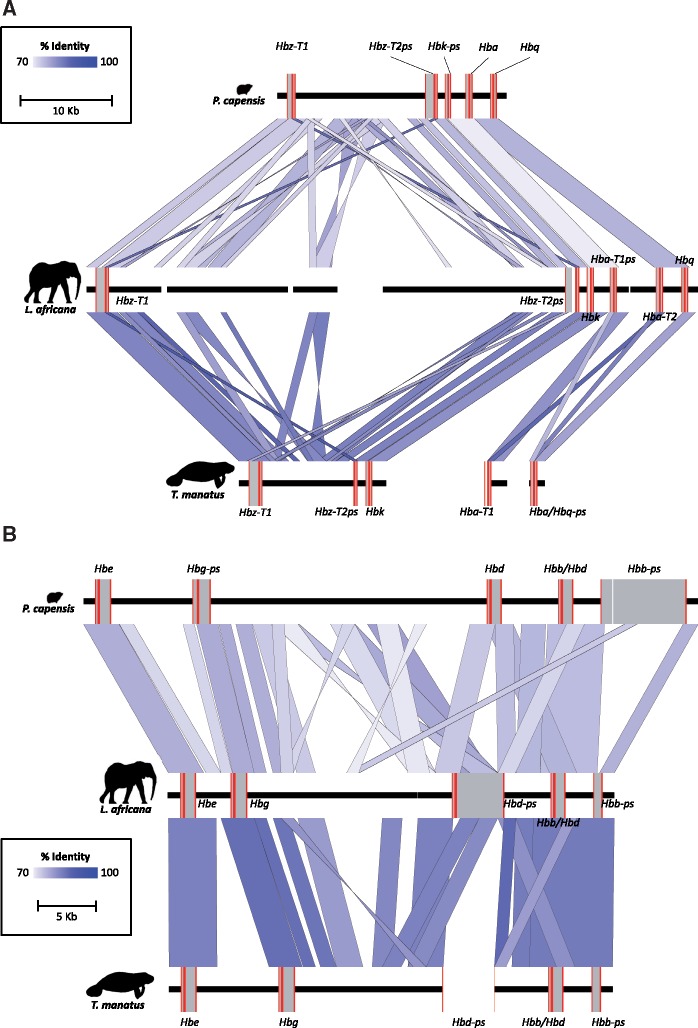
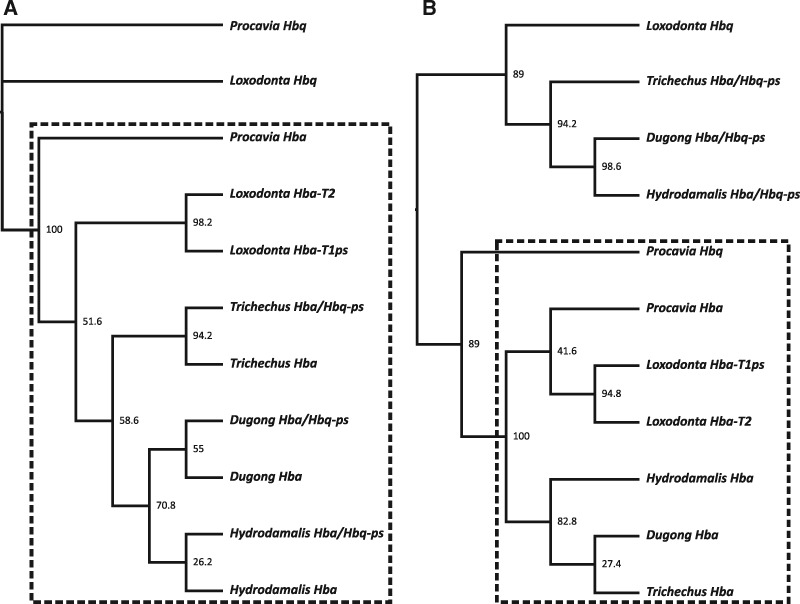
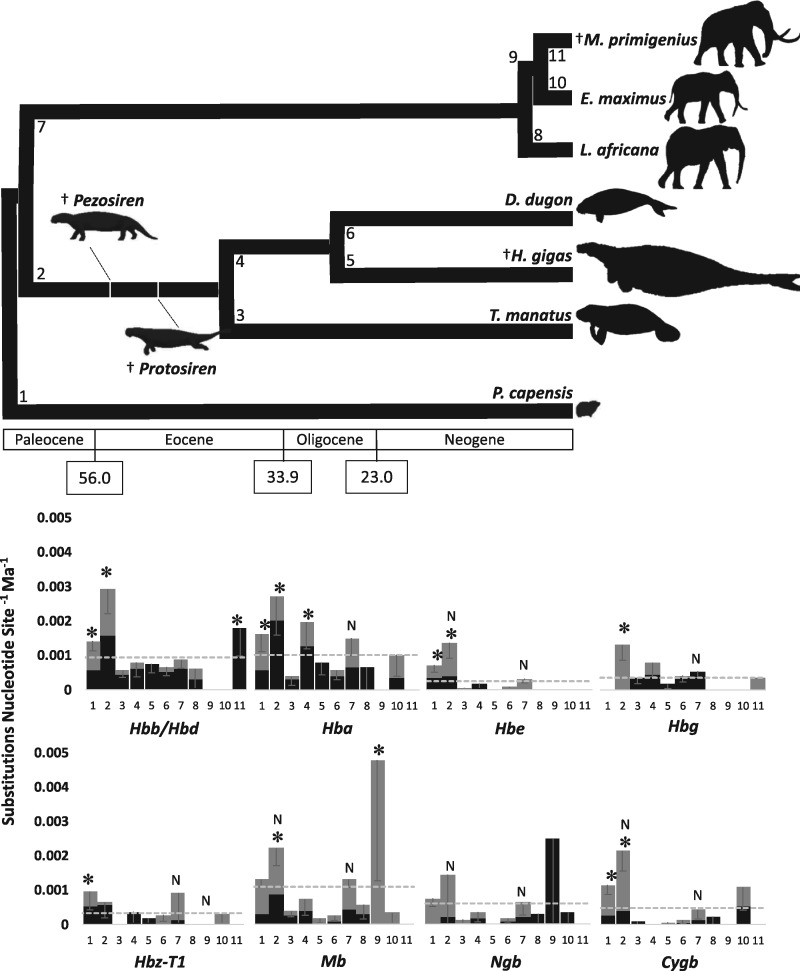
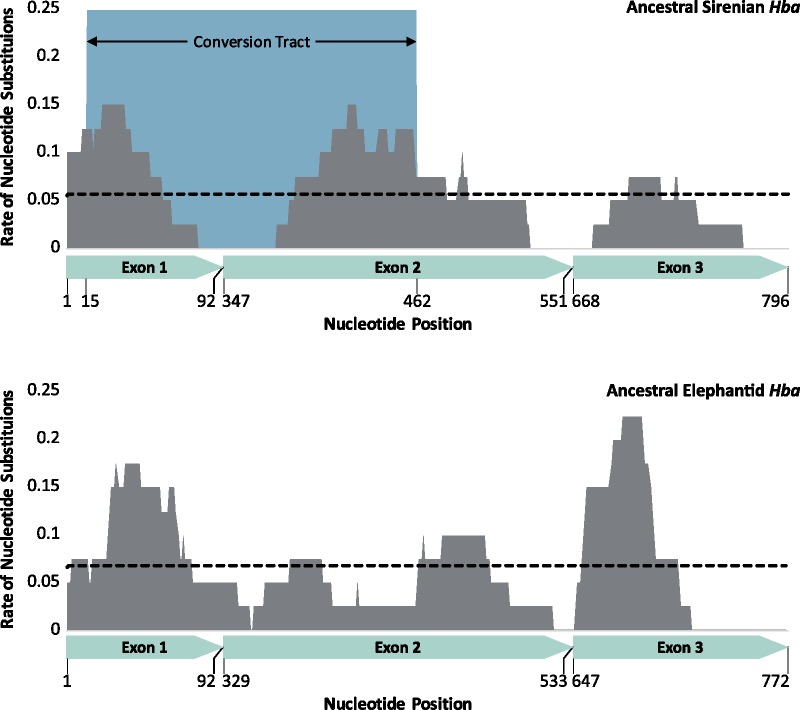
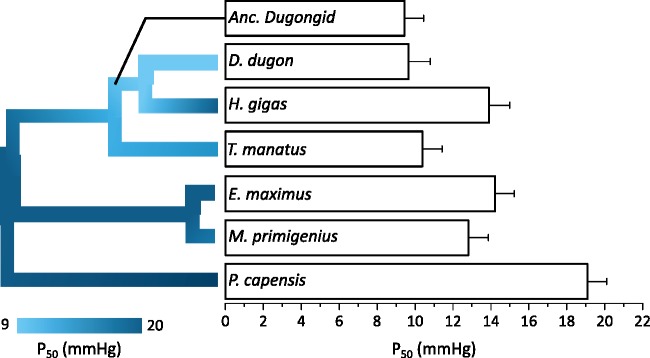
As limits on O2 availability during submergence impose severe constraints on aerobic respiration, the oxygen binding globin proteins of marine mammals are expected to have evolved under strong evolutionary pressures during their land-to-sea transition. Here, we address this question for the order Sirenia by retrieving, annotating, and performing detailed selection analyses on the globin repertoire of the extinct Steller's sea cow (Hydrodamalis gigas), dugong (Dugong dugon), and Florida manatee (Trichechus manatus latirostris) in relation to their closest living terrestrial relatives (elephants and hyraxes). These analyses indicate most loci experienced elevated nucleotide substitution rates during their transition to a fully aquatic lifestyle. While most of these genes evolved under neutrality or strong purifying selection, the rate of nonsynonymous/synonymous replacements increased in two genes (Hbz-T1 and Hba-T1) that encode the α-type chains of hemoglobin (Hb) during each stage of life. Notably, the relaxed evolution of Hba-T1 is temporally coupled with the emergence of a chimeric pseudogene (Hba-T2/Hbq-ps) that contributed to the tandemly linked Hba-T1 of stem sirenians via interparalog gene conversion. Functional tests on recombinant Hb proteins from extant and ancestral sirenians further revealed that the molecular remodeling of Hba-T1 coincided with increased Hb-O2 affinity in early sirenians. Available evidence suggests that this trait evolved to maximize O2 extraction from finite lung stores and suppress tissue O2 offloading, thereby facilitating the low metabolic intensities of extant sirenians. In contrast, the derived reduction in Hb-O2 affinity in (sub)Arctic Steller's sea cows is consistent with fueling increased thermogenesis by these once colossal marine herbivores.
Keywords: ancient DNA; aquatic adaptation; cytoglobin; gene conversion; hemoglobin; molecular evolution; myoglobin; neuroglobin; oxygen affinity; pseudogene.
© The Author(s) 2019. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures
References
-
- Benjamini Y, Hochberg Y.. 1995. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc B. 57:289–300.
-
- Blessing MH. 1972. Studies on the concentration of myoglobin in the sea-cow and porpoise. Comp Biochem Physiol A. 413:475–480. - PubMed
-
- Bonaventura J, Bonaventura C, Sullivan B, Ferruzzi G, McCurdy PR, Fox J, Moo-Penn WF.. 1976. Hemoglobin providence. Functional consequences of two alterations of the 2, 3-diphosphoglycerate binding site at position beta 82. J Biol Chem. 25123:7563–7571. - PubMed
-
- Burmester T, Haberkamp M, Mitz S, Roesner A, Schmidt M, Ebner B, Gerlach F, Fuchs C, Hankeln T.. 2004. Neuroglobin and cytoglobin: genes, proteins and evolution. IUBMB Life 56(11–12):703–707. - PubMed
-
- Burmester T, Hankeln T.. 2014. Function and evolution of vertebrate globins. Acta Physiol. 2113:501–514. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
