

Nanoneedle-Mediated Stimulation of Cell Mechanotransduction Machinery
- PMID: 30829469
- PMCID: PMC6439438
- DOI: 10.1021/acsnano.8b06998
Nanoneedle-Mediated Stimulation of Cell Mechanotransduction Machinery
Abstract
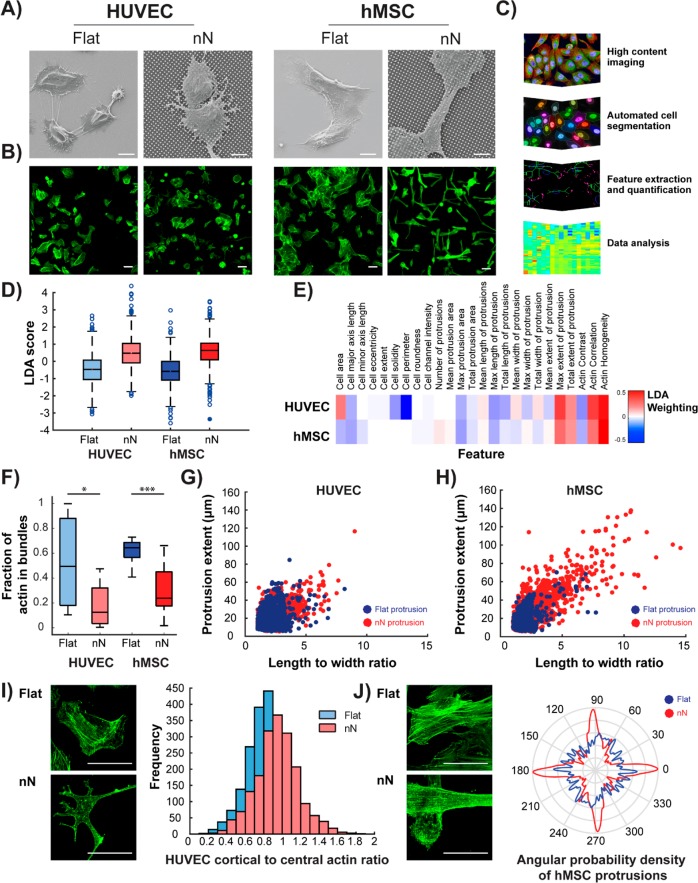
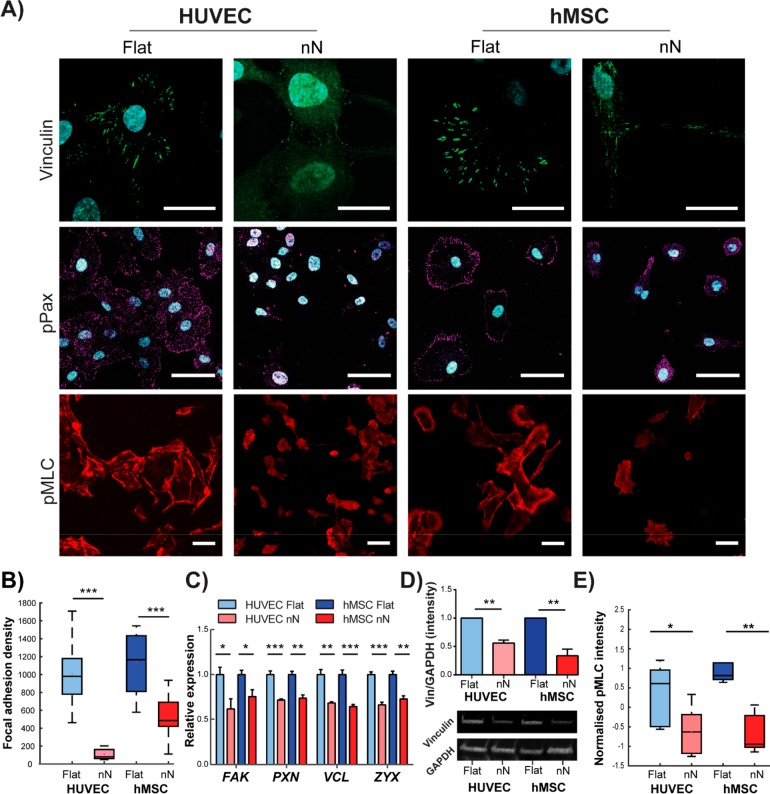
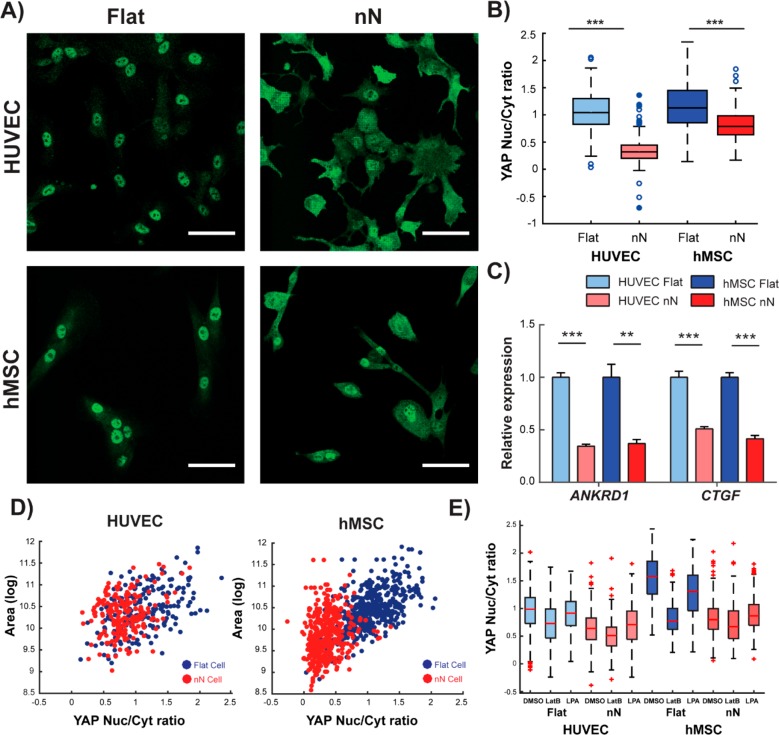
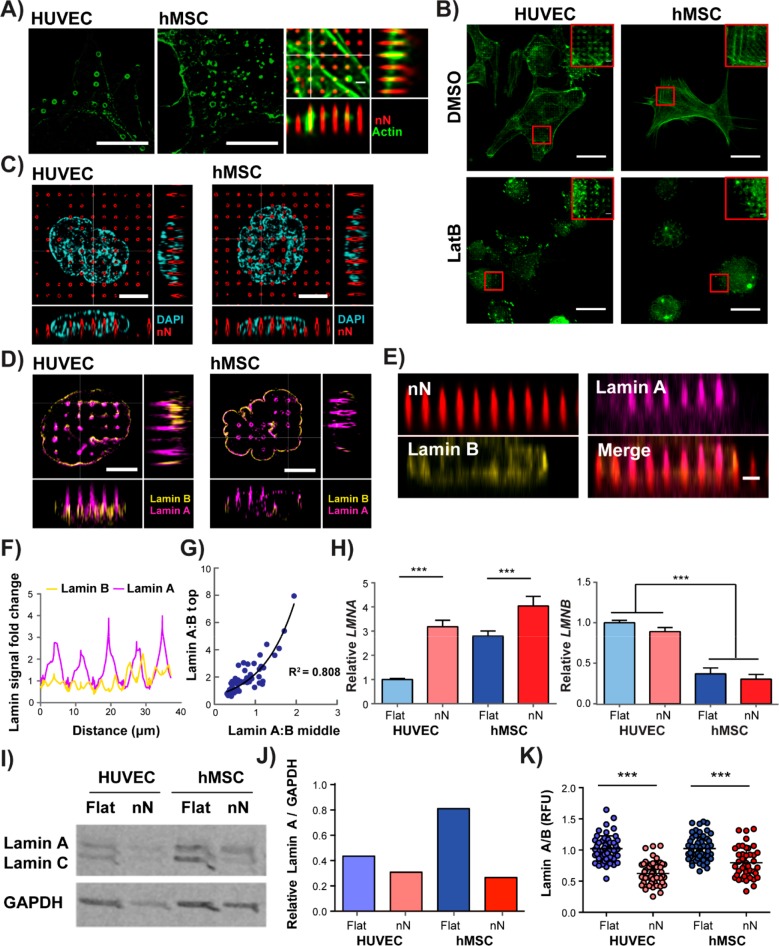
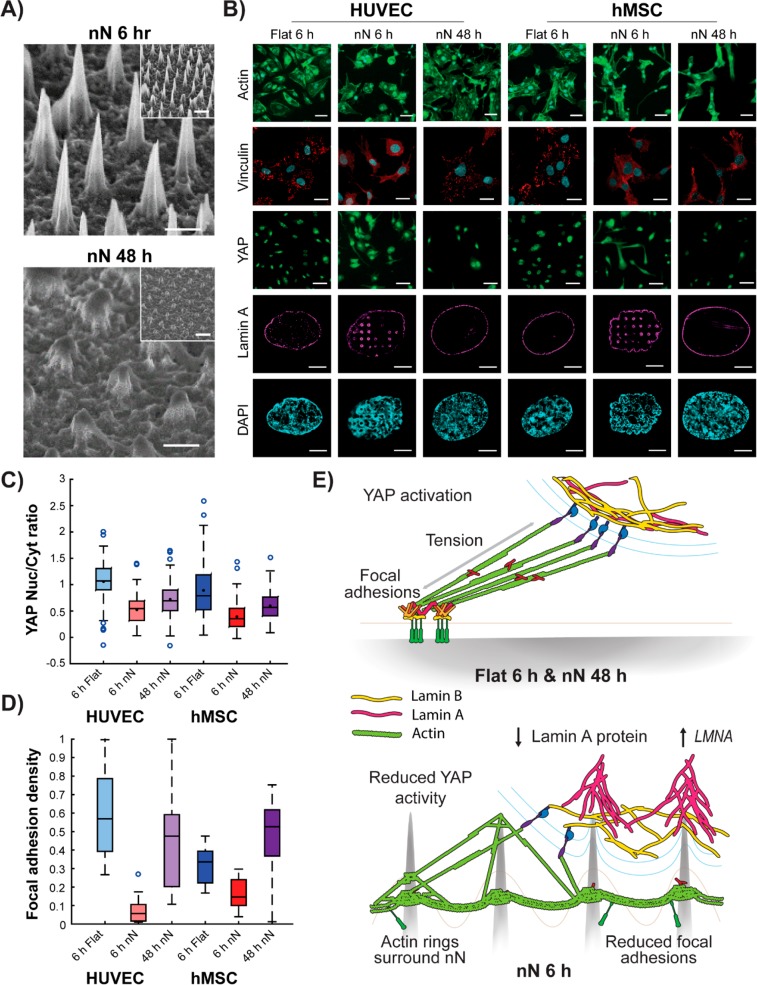
Biomaterial substrates can be engineered to present topographical signals to cells which, through interactions between the material and active components of the cell membrane, regulate key cellular processes and guide cell fate decisions. However, targeting mechanoresponsive elements that reside within the intracellular domain is a concept that has only recently emerged. Here, we show that mesoporous silicon nanoneedle arrays interact simultaneously with the cell membrane, cytoskeleton, and nucleus of primary human cells, generating distinct responses at each of these cellular compartments. Specifically, nanoneedles inhibit focal adhesion maturation at the membrane, reduce tension in the cytoskeleton, and lead to remodeling of the nuclear envelope at sites of impingement. The combined changes in actin cytoskeleton assembly, expression and segregation of the nuclear lamina, and localization of Yes-associated protein (YAP) correlate differently from what is canonically observed upon stimulation at the cell membrane, revealing that biophysical cues directed to the intracellular space can generate heretofore unobserved mechanosensory responses. These findings highlight the ability of nanoneedles to study and direct the phenotype of large cell populations simultaneously, through biophysical interactions with multiple mechanoresponsive components.
Keywords: cell−material interactions; mechanotransduction; nanoneedles; nuclear mechanics; porous silicon; super-resolution microscopy.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
-
- McMurray R. J.; Gadegaard N.; Tsimbouri P. M.; Burgess K. V.; McNamara L. E.; Tare R.; Murawski K.; Kingham E.; Oreffo R. O. C.; Dalby M. J. Nanoscale Surfaces for the Long-Term Maintenance of Mesenchymal Stem Cell Phenotype and Multipotency. Nat. Mater. 2011, 10, 637–644. 10.1038/nmat3058. - DOI - PubMed
-
- Swift J.; Ivanovska I. L.; Buxboim A.; Harada T.; Dingal P. C. D. P.; Pinter J.; Pajerowski J. D.; Spinler K. R.; Shin J. W.; Tewari M.; Rehfeldt F.; Speicher D. W.; Discher D. E. Nuclear Lamin-A Scales with Tissue Stiffness and Enhances Matrix-Directed Differentiation. Science 2013, 341, 1240104.10.1126/science.1240104. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- NC/R001006/1/NC3RS_/National Centre for the Replacement, Refinement and Reduction of Animals in Research/United Kingdom
- BB/L015129/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- 20146/CRUK_/Cancer Research UK/United Kingdom
- WT_/Wellcome Trust/United Kingdom
- C37275/1A20146/CRUK_/Cancer Research UK/United Kingdom
