

Ash1 counteracts Polycomb repression independent of histone H3 lysine 36 methylation
- PMID: 30833342
- PMCID: PMC6446198
- DOI: 10.15252/embr.201846762
Ash1 counteracts Polycomb repression independent of histone H3 lysine 36 methylation
Abstract
Polycomb repression is critical for metazoan development. Equally important but less studied is the Trithorax system, which safeguards Polycomb target genes from the repression in cells where they have to remain active. It was proposed that the Trithorax system acts via methylation of histone H3 at lysine 4 and lysine 36 (H3K36), thereby inhibiting histone methyltransferase activity of the Polycomb complexes. Here we test this hypothesis by asking whether the Trithorax group protein Ash1 requires H3K36 methylation to counteract Polycomb repression. We show that Ash1 is the only Drosophila H3K36-specific methyltransferase necessary to prevent excessive Polycomb repression of homeotic genes. Unexpectedly, our experiments reveal no correlation between the extent of H3K36 methylation and the resistance to Polycomb repression. Furthermore, we find that complete substitution of the zygotic histone H3 with a variant in which lysine 36 is replaced by arginine does not cause excessive repression of homeotic genes. Our results suggest that the model, where the Trithorax group proteins methylate histone H3 to inhibit the histone methyltransferase activity of the Polycomb complexes, needs revision.
Keywords: Drosophila; Ash1; H3K36 methylation; Polycomb; Trithorax.
© 2019 The Authors.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
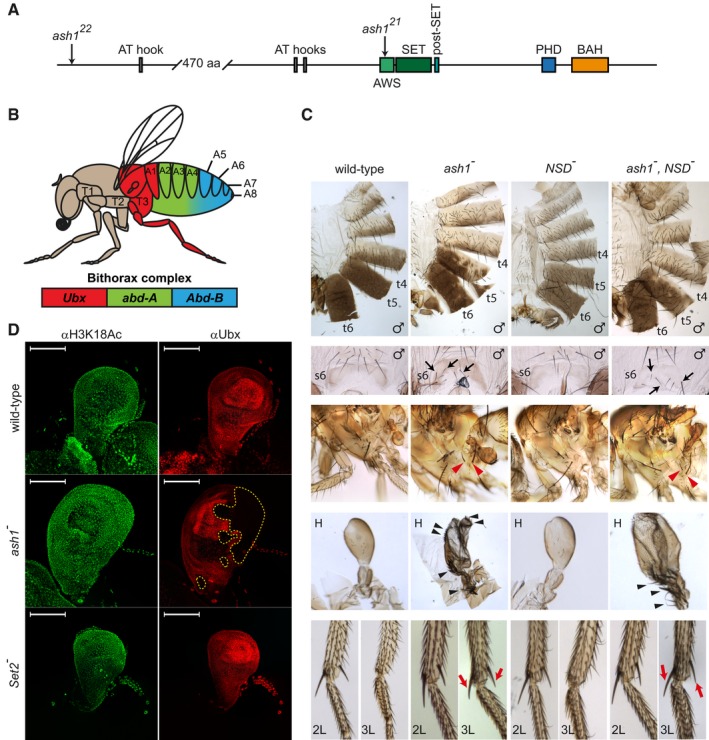
The schematic of the Drosophila Ash1 protein organization. Ash1 is 2,226 amino acid long and contains eight domains (indicated by coloured rectangles). The SET domain together with the AWS (Associated With SET) and the post‐SET domains are necessary and sufficient for Ash1 histone methyltransferase (HMTase) activity. The functions of the BAH (Bromo Adjacent Homology), PHD (Plant homeodomain) and AT‐hook domains are unknown. The positions of ash1 22 and ash1 21 point mutations are indicated by arrows.
Segmental expression of the Drosophila bithorax complex genes. The three genes of the complex, Ubx, abd‐A and Abd‐B, are shown as coloured rectangles. The expression of Ubx gives identity to the third thoracic (T3) and the first abdominal (A1) segments, the expression of abd‐A defines the second, third and fourth abdominal segments (A2–A4), and the expression of Abd‐B gives identity to the rest of the abdominal segments (illustrated with corresponding colour code).
Adult phenotypes of the ash1 and NSD mutants. In ash1 22/ash1 21 mutants (designated as ash1 −), the loss of Abd‐B expression results in partial transformation of abdominal segments 6 and 5 towards segments 5 and 4, which is visible from the partial loss of pigmentation on tergites 5 and 6 (t5 and t6) and appearance of bristles on sternite 6 (s6, marked with black arrows). The loss of Ubx expression causes transformation of the third thoracic to the second thoracic segment visible as partial haltere (H) to wing and third leg (3L) to second leg (2L) transformations. The former is evident from the change in the haltere shape and the appearance of multiple bristles (black arrowheads). The latter is indicated by the apical and pre‐apical bristles (red arrows) on the tibia of the third leg of ash1 mutants. These are normally present on 2L but absent on 3L (compare to wild‐type). Also note the appearance of additional hypopleural bristles on the third thoracic segment of the ash1 − flies (red arrowheads), which indicate its transformation towards the second thoracic segment. Phenotype of the NSD ds46/NSD ds46 (NSD −) flies is indistinguishable from wild‐type and the phenotype of the double ash1 22 ,NSD ds46/ash1 22 ,NSD ds46 (ash1 − ,NSD −) flies is no more severe than that of the single ash1 22/ash1 21 (ash1 −) mutants.
Ubx expression in the haltere imaginal discs. The expression was assayed by immunostaining with antibodies against Ubx (red) and acetylated H3K18 (green, positive control). While ash1 22/ash1 21 (ash1 −) larvae show stochastic clonal loss of the Ubx immunostaining in haltere discs (yellow dashed lines), Set2 − larvae have uniform expression of Ubx throughout the haltere disc, resembling that in the wild‐type larvae. Scale bars indicate 100 μm.
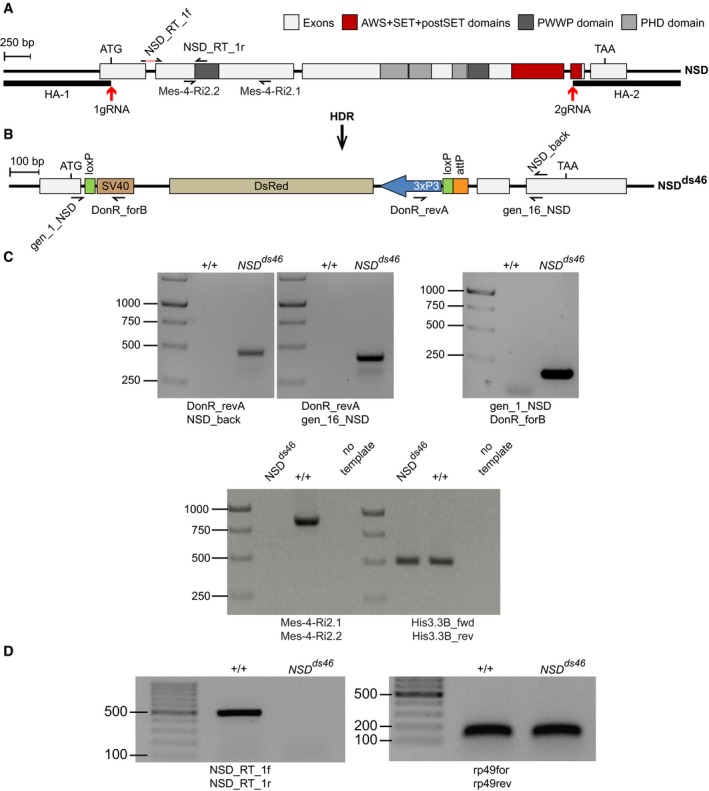
The structure of the Drosophila NSD locus. Red arrows indicate locations of the gRNA sites used for CRISPR/Cas9‐mediated replacement of the NSD Open Reading Frame (ORF) with DsRed. The homology regions HA‐1 and HA‐2, used for the replacement, are shown with bold lines. The half‐arrows represent the primers used for genotyping of the mutant allele. The dashed red line in NSD_RT_1f primer, used for RT–qPCR, indicates the intronic region that is excluded from the primer.
The NSD ds46 allele. After Homology‐directed Repair (HDR), the insertion of the DsRed cassette generates the loss‐of‐function allele NSD ds46, where DsRed substitutes most of the NSD ORF. The DsRed expression is controlled by the 3xP3 promoter. In addition to DsRed, the replacement cassette contains two loxP sites to remove DsRed via Cre‐mediated recombination and an attP docking site to insert variants of the NSD ORF. The half‐arrows represent the primers used to genotype the mutant allele.
Genotyping of the NSD ds46 allele by PCR. The replacement of the NSD ORF by the DsRed cassette was confirmed by PCR with four different primers pairs. Three primer pairs (top row of images) yield the product only if the replacement has happened. The expected sizes of the PCR products are 469, 427 and 186 bp for DonR_revA and NSD_back, DonR_revA and gen_16_NSD, and gene_1_NSD and DonR_forB primer pairs, respectively. The PCR with Mes‐4‐RI‐2.1 and Mes‐4‐RI‐2.2 primer pair amplifies the 830 bp product only from the wild‐type allele. PCR with His3.3B_fwd and His3.3B_rev primer pair, amplifying the 495 bp DNA fragment from the His3.3B gene, was used as a positive control.
The NSD ds46 allele produces no messenger RNA. This was confirmed by RT–PCR with NSD_Rt_1f and NSD_Rt_1r primer pair, which yields the 513 bp product only when the intact NSD mRNA is produced. RT–PCR with rp49for and rp49rev primer pair that amplifies 132 bp fragment from the cDNA of constitutively expressed RpL32 (a.k.a rp49) gene was used as positive control.
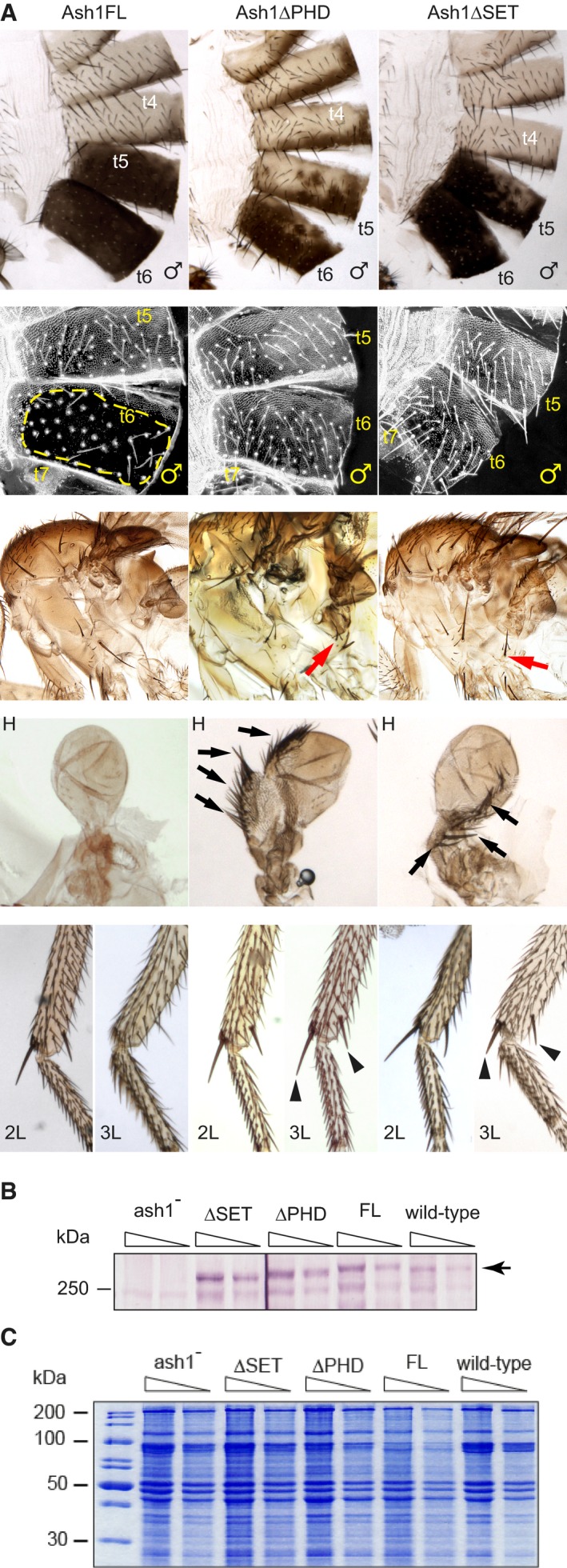
Adult phenotypes of the ash1 22/ash1 9011 flies supplemented with transgenic constructs expressing either the full‐length Ash1 (Ash1FL) or the truncated variants lacking the PHD (Ash1ΔPHD) or the SET (Ash1ΔSET) domains. Note extra hypopleural bristles (red arrows), the third leg (L3) to second leg (L2), haltere (H) to wing, t5–t4 and t6–t5 transformations in the Ash1ΔPHD and the Ash1ΔSET but not in the Ash1FL flies. The latter are evident from the partial loss of pigmentation in t6 and t5, or the appearance of small bristles (trichomas) on t6 of the Ash1ΔPHD and the Ash1∆SET flies in the area that is normally naked (Ash1FL, yellow dashed line). The transformed L3 acquire apical and pre‐apical bristles on the tibia (black triangles) while halteres change shape and acquire rows of bristles (black arrows).
Twofold dilutions of total nuclear protein extracts from the third instar larvae of the ash1 22/ash1 9011 mutants supplemented with various transgenic constructs and wild‐type flies were analysed by Western blot with anti‐Ash1 antibodies. Arrow indicates the position of Ash1 protein. Note that transgenic proteins are expressed at comparable level.
Coomassie staining of SDS–PAGE separated protein extracts from (B) was used to control the loading.
- A–C
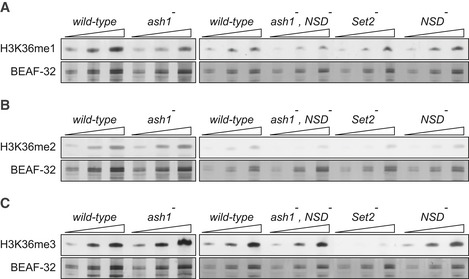
Twofold serial dilutions of the total protein extracts from the wild‐type, ash1 22/ash1 9011 (ash1 −), ash1 22 ,NSD ds46/ash1 9011 ,NSD ds46 (ash1 − , NSD −) and Set2 1 (Set2 −) larval brains, imaginal discs and salivary glands were analysed by Western blot with antibodies against H3K36me1 (A), H3K36me2 (B) and H3K36me3 (C). Note the strong (> 10‐fold) reduction of H3K36me3 signal in the Set2 − extract and the slight (˜ 2‐fold) reduction of H3K36me1 signal in the ash1 − and ash1 − , NSD − extracts. The protein extracts from the wild‐type, double ash1 − , NSD − and single NSD − and Set2 − mutants (right panels) were analysed together on the same membrane; however, the images of the H3K36me1 and H3K36me3 Western blots were modified to splice out the marker lane between the ash1 − , NSD − and the Set2 − extracts. Western blots with constitutively expressed BEAF‐32 protein were used as loading controls.
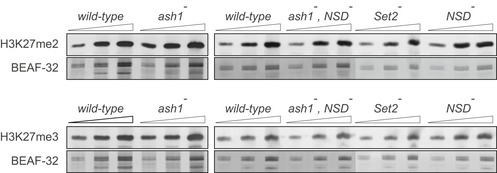
- A–C
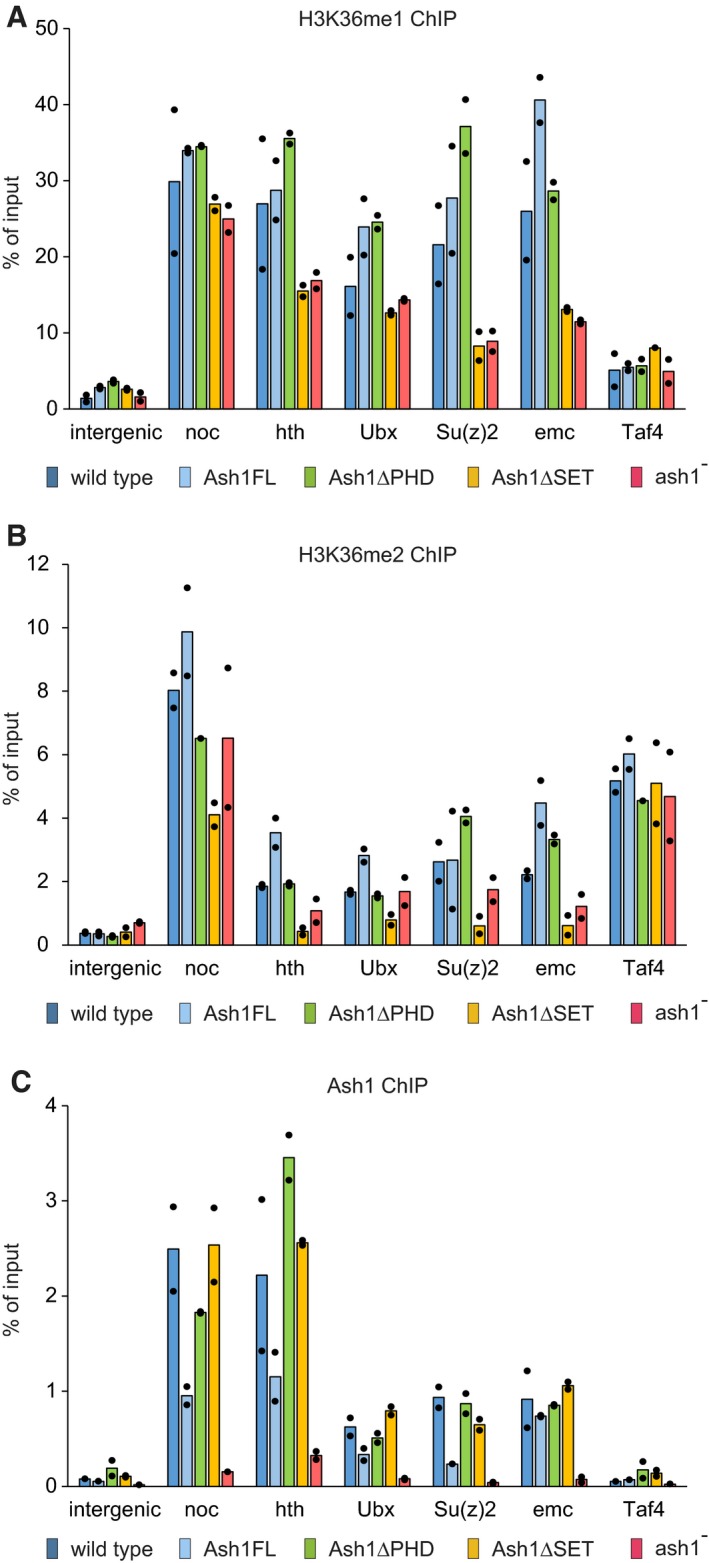
Chromatin from the wild‐type (dark blue bars), ash1 22/ash1 9011 (ash1−, red bars) and transgenic ash1 22/ash1 9011 (Ash1FL, light blue bars; Ash1ΔPHD, green bars; Ash1ΔSET, orange bars) larvae was subjected to immunoprecipitation with the antibodies against H3K36me1 (A), H3K36me2 (B) and Ash1 (C). Histograms show the mean of the two independent experiments (n = 2) with dots indicating individual experimental results. An intergenic region on chromosome 3R (intergenic) and the constitutively active TBP‐associated factor 4 (Taf4) gene serve as controls. The loss of Ash1 ChIP signal in the ash1 − larvae indicates that the selected genes are the genuine Ash1 binding sites.
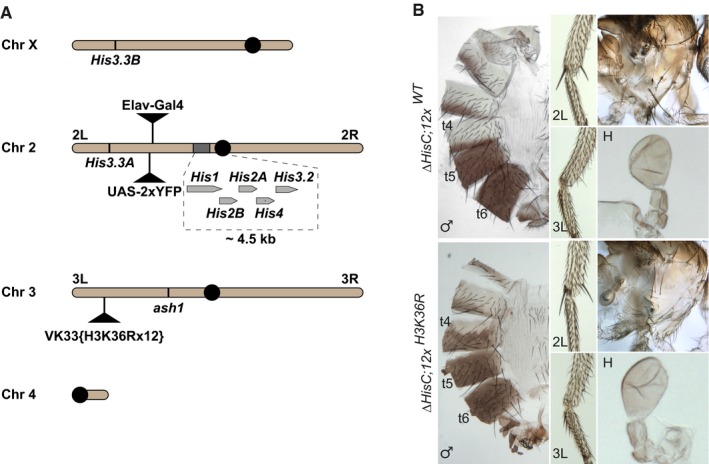
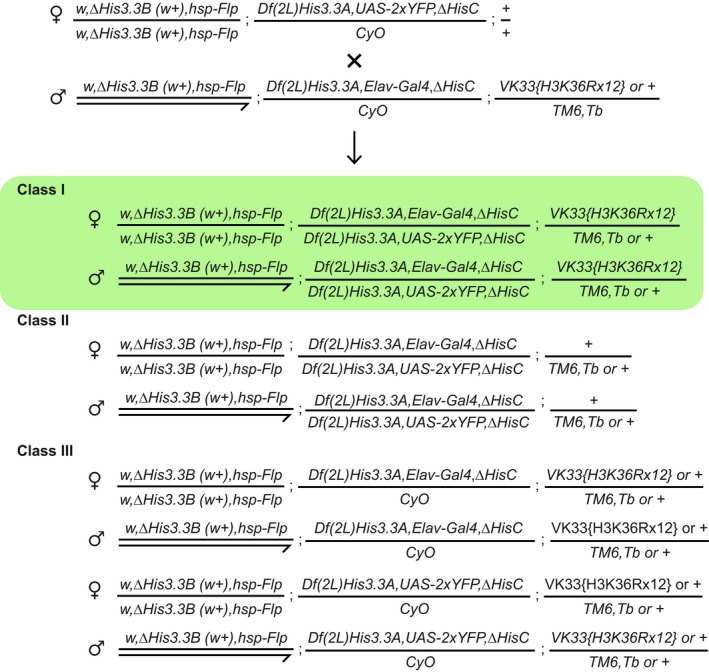
Chromosomal positions of various histone H3 genes, ash1 and marker transgenes. Twenty‐three histone gene repeat units, each containing single His1, His2B, His2A, His4 and His3.2 gene, are clustered near the centromere (black circle) of chromosome 2. These histone genes are removed by the ΔHisC deletion. To select the animals homozygous for the deletion, the ΔHisC chromosomes are marked with insertions of either Elav‐Gal4 or UAS‐2xYFP transgenes (black triangles on chromosome 2L). The His3.3A and His3.3B genes encode the same protein, but reside on chromosomes 2L and X. The transgenes carrying twelve copies of either the wild‐type histone repeat unit (12x WT) or the unit in which H3 gene is altered to have K36 replaced with arginine (12x H3K36R) are inserted in the same attP site (black triangle on chromosome 3L) 47. The ash1 gene is located on the same chromosome arm.
The ΔHisC; 12x H3K36R and control ΔHisC; 12x WT flies show no homeotic transformations and are indistinguishable from the wild type.
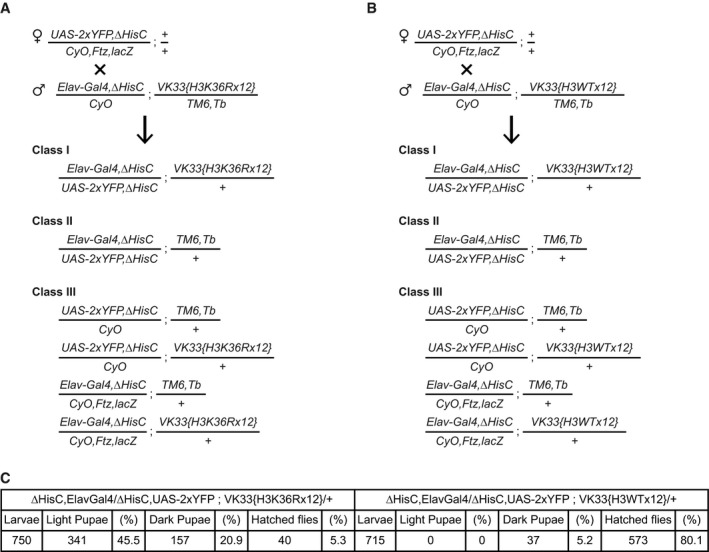
- A, B
To generate the desired flies, females carrying ΔHisC recombined with the UAS‐2xYFP transgene were crossed to males carrying ΔHisC recombined with the Elav‐Gal4 transgene and one of the two variants of the transgene with 12 copies of the histone gene cluster. In one of the transgenes, all copies of His3.2 gene contained H3K36R mutation (A) and in the other, all copies of His3.2 were wild‐type (B). The crossing scheme and the progeny classes in (A) and (B) were identical. The Class I larvae were separated from the larvae of the other classes, based on the YFP signal. Although the Class II animals also show YFP signal, they die at early embryonic stages after running out of maternally deposited histones. The Class III larvae lack YFP signal and are easily distinguishable from Class I.
- C
Twelve transgenic copies of the wild‐type histone gene cluster complement the loss of endogenous HisC locus nearly completely (80.1%). However, the transgene carrying H3K36R mutation fails to compensate for the deletion of the endogenous histones cluster. Although a large fraction of these animals reaches the pupal stage (about 66%), only 5% of them reach the adult stage.
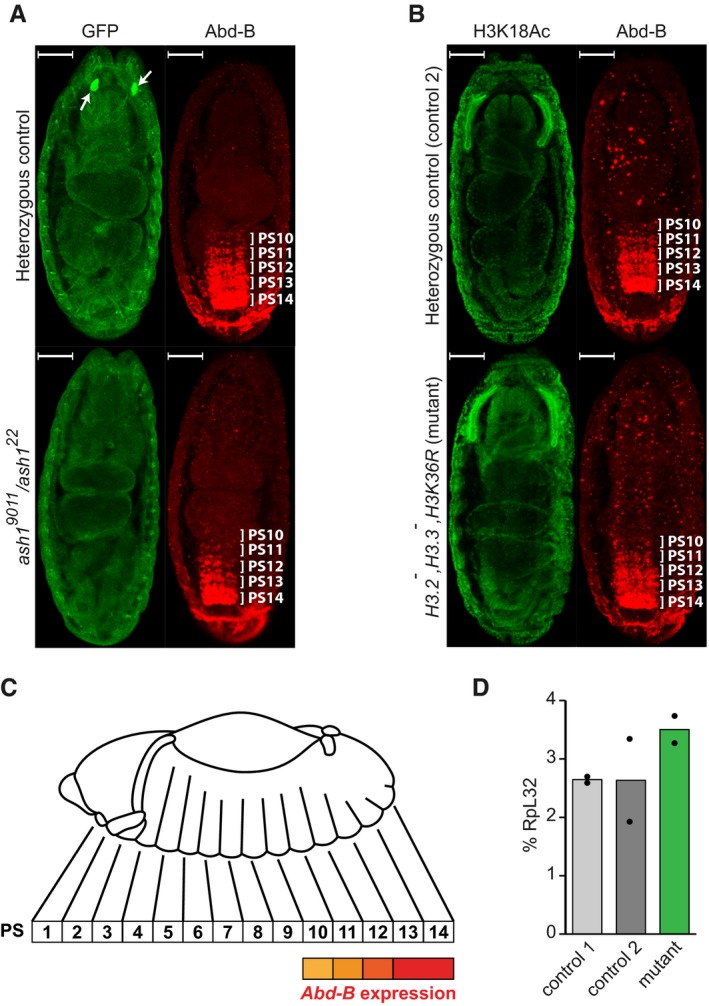
The Abd‐B expression in the Central Nervous System (CNS) of stage 16 embryos was assayed by immunostaining with antibodies against Abd‐B (red). In the ash1 22/TM3, Sb, e Kr::GFP or ash1 9011/TM3, Sb, e Kr::GFP embryos (heterozygous control), Abd‐B is expressed in parasegments 14–10 in a gradient that recedes from the posterior to anterior parasegment. In the ash1 22/ash1 9011 mutants, the Abd‐B gradient is much steeper, with reduced staining of parasegments 13 and 12 and at the edge of the detection in parasegments 11–10. Heterozygous control and ash1 22/ash1 9011 mutant embryos were stained together and separated by strong GFP expression (green) in the Bolwig's organs (white arrows). Here and in (B) the embryos are oriented with anterior pole facing the top and scaling bars correspond to 50 μm.
Unlike ash1 mutants, embryos homozygous for His3.3A, His3.3B, ΔHisC deletions and supplemented with 12x H3K36R transgene (His3.2 − ,His3.3 − ,H3K36R) display the wild‐type Abd‐B immunostaining pattern (red), the same as the control embryos heterozygous for His3.3A, and ΔHisC deletions. In this experiment, immunostaining with antibodies against acetylated H3K18 (green) served as positive control.
The schematic of the Abd‐B expression in the Drosophila embryo. Although homeotic transformations are often described according to the adult segments affected, the expression of the bithorax complex genes is regulated at the level of embryonic parasegments (PS), which are directly related to adult segments but slightly shifted relative to one another 5, 64.
Reverse transcription and quantitative PCR (RT–qPCR) measurement of Abd‐B expression in His3.2 − ,His3.3 − ,H3K36R embryos. Expression of Abd‐B in stage 16 embryos, which are homozygous for His3.3A, His3.3B and ∆HisC deletions and carry 12x H3K36R transgene (mutant), is not reduced compared to their wild‐type counterparts (control 1) or embryos heterozygous for His3.3A, and ΔHisC deletions (control 2). Histograms show the mean of the two independent experiments (n = 2) with dots indicating individual experimental results.
References
-
- Schuettengruber B, Bourbon HM, Di Croce L, Cavalli G (2017) Genome regulation by Polycomb and Trithorax: 70 years and counting. Cell 171: 34–57 - PubMed
-
- Schwartz YB, Pirrotta V (2007) Polycomb silencing mechanisms and the management of genomic programmes. Nat Rev Genet 8: 9–22 - PubMed
-
- Lewis EB (1978) A gene complex controlling segmentation in Drosophila . Nature 276: 565–570 - PubMed
-
- Maeda RK, Karch F (2006) The ABC of the BX‐C: the bithorax complex explained. Development 133: 1413–1422 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
