

The Proteasome and Its Network: Engineering for Adaptability
- PMID: 30833452
- PMCID: PMC6829053
- DOI: 10.1101/cshperspect.a033985
The Proteasome and Its Network: Engineering for Adaptability
Abstract
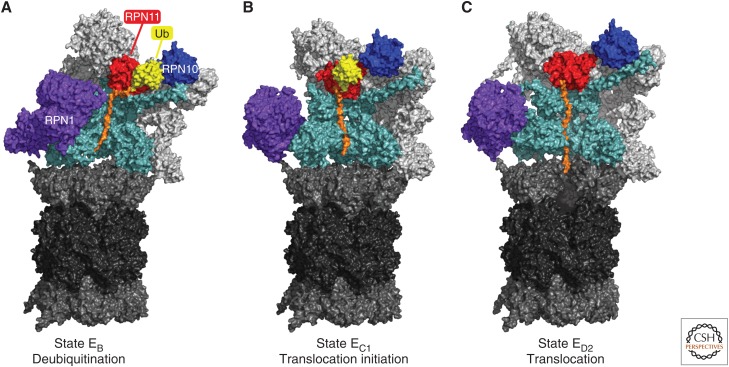
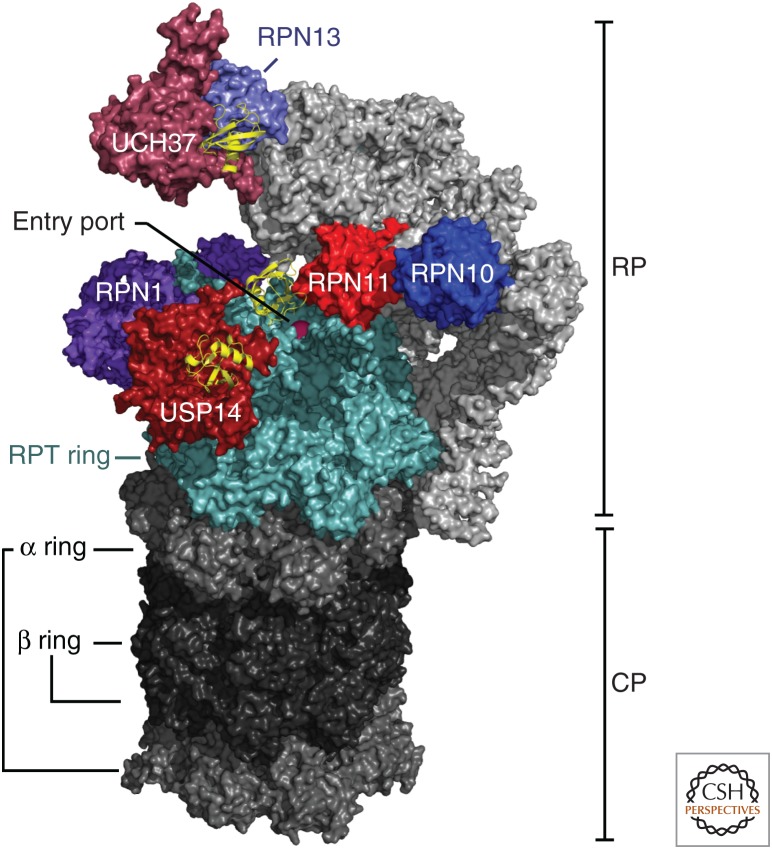
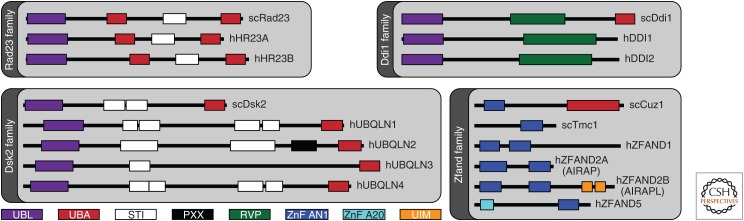
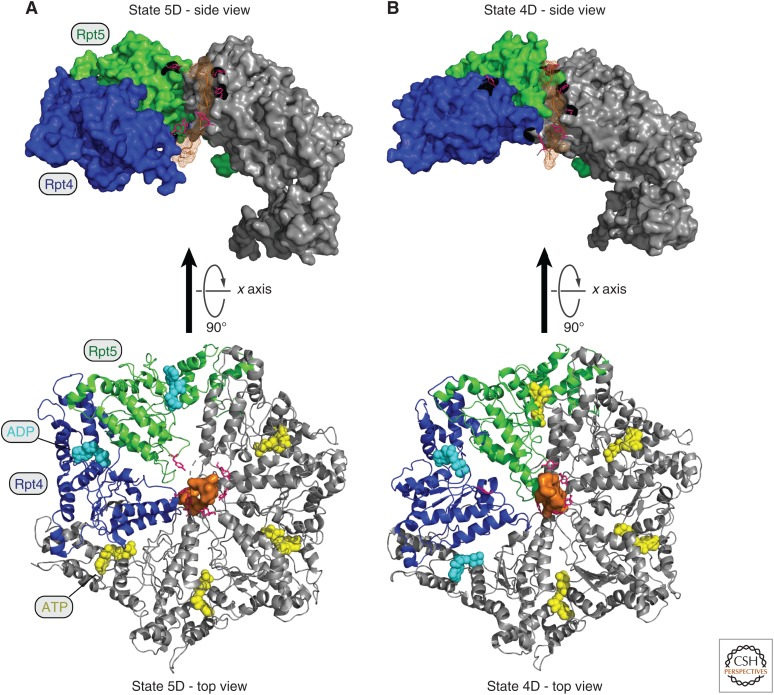
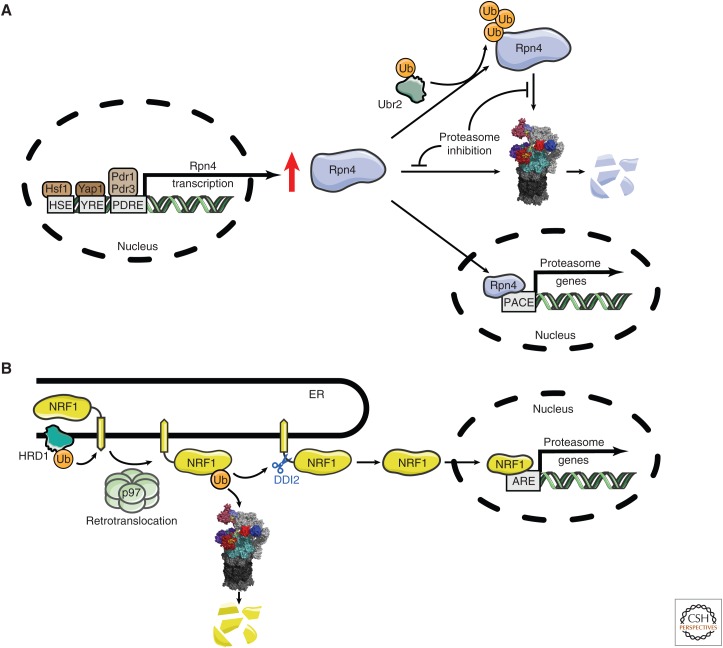
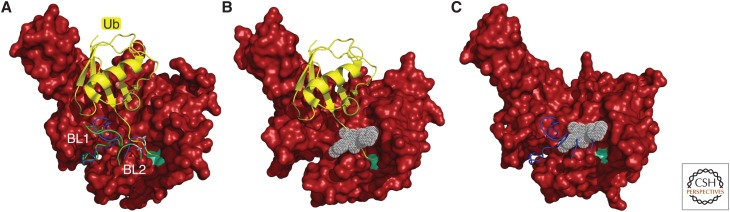
The proteasome, the most complex protease known, degrades proteins that have been conjugated to ubiquitin. It faces the unique challenge of acting enzymatically on hundreds and perhaps thousands of structurally diverse substrates, mechanically unfolding them from their native state and translocating them vectorially from one specialized compartment of the enzyme to another. Moreover, substrates are modified by ubiquitin in myriad configurations of chains. The many unusual design features of the proteasome may have evolved in part to endow this enzyme with a robust ability to process substrates regardless of their identity. The proteasome plays a major role in preserving protein homeostasis in the cell, which requires adaptation to a wide variety of stress conditions. Modulation of proteasome function is achieved through a large network of proteins that interact with it dynamically, modify it enzymatically, or fine-tune its levels. The resulting adaptability of the proteasome, which is unique among proteases, enables cells to control the output of the ubiquitin-proteasome pathway on a global scale.
Copyright © 2020 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources