

Functional Modules of the Proteostasis Network
- PMID: 30833457
- PMCID: PMC6942124
- DOI: 10.1101/cshperspect.a033951
Functional Modules of the Proteostasis Network
Abstract
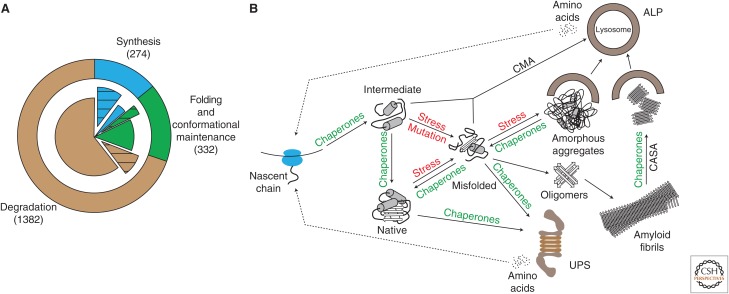
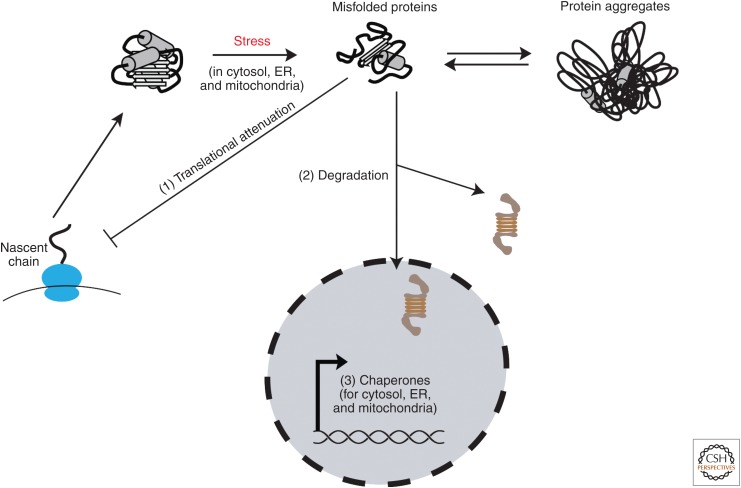
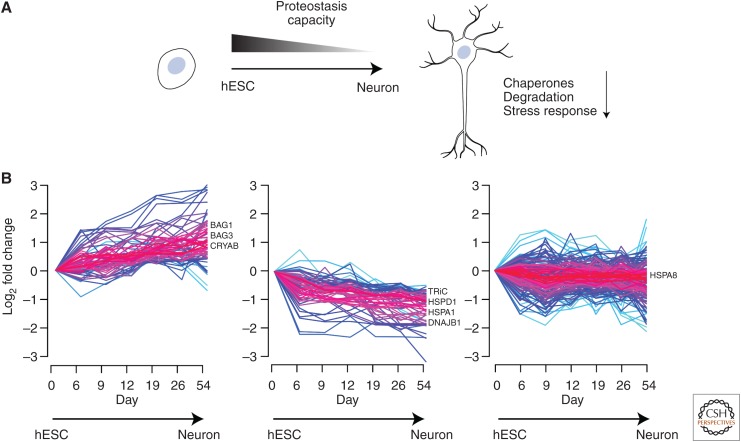
Cells invest in an extensive network of factors to maintain protein homeostasis (proteostasis) and prevent the accumulation of potentially toxic protein aggregates. This proteostasis network (PN) comprises the machineries for the biogenesis, folding, conformational maintenance, and degradation of proteins with molecular chaperones as central coordinators. Here, we review recent progress in understanding the modular architecture of the PN in mammalian cells and how it is modified during cell differentiation. We discuss the capacity and limitations of the PN in maintaining proteome integrity in the face of proteotoxic stresses, such as aggregate formation in neurodegenerative diseases. Finally, we outline various pharmacological interventions to ameliorate proteostasis imbalance.
Copyright © 2020 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical