

Systems Analysis of the 22q11.2 Microdeletion Syndrome Converges on a Mitochondrial Interactome Necessary for Synapse Function and Behavior
- PMID: 30833507
- PMCID: PMC6495129
- DOI: 10.1523/JNEUROSCI.1983-18.2019
Systems Analysis of the 22q11.2 Microdeletion Syndrome Converges on a Mitochondrial Interactome Necessary for Synapse Function and Behavior
Abstract
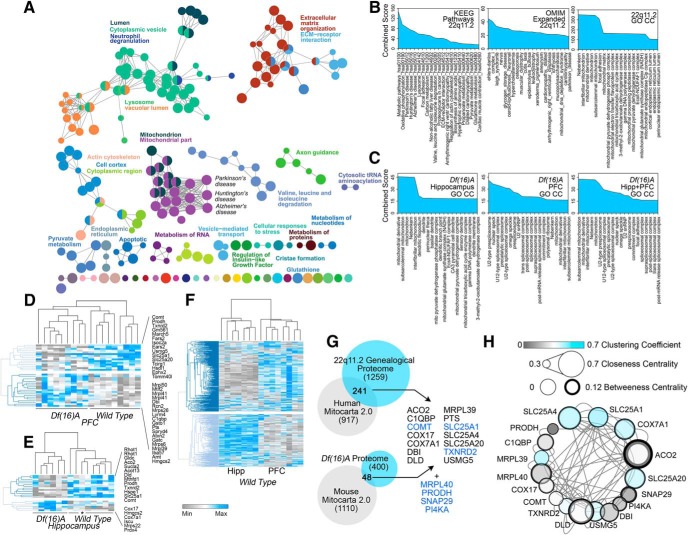
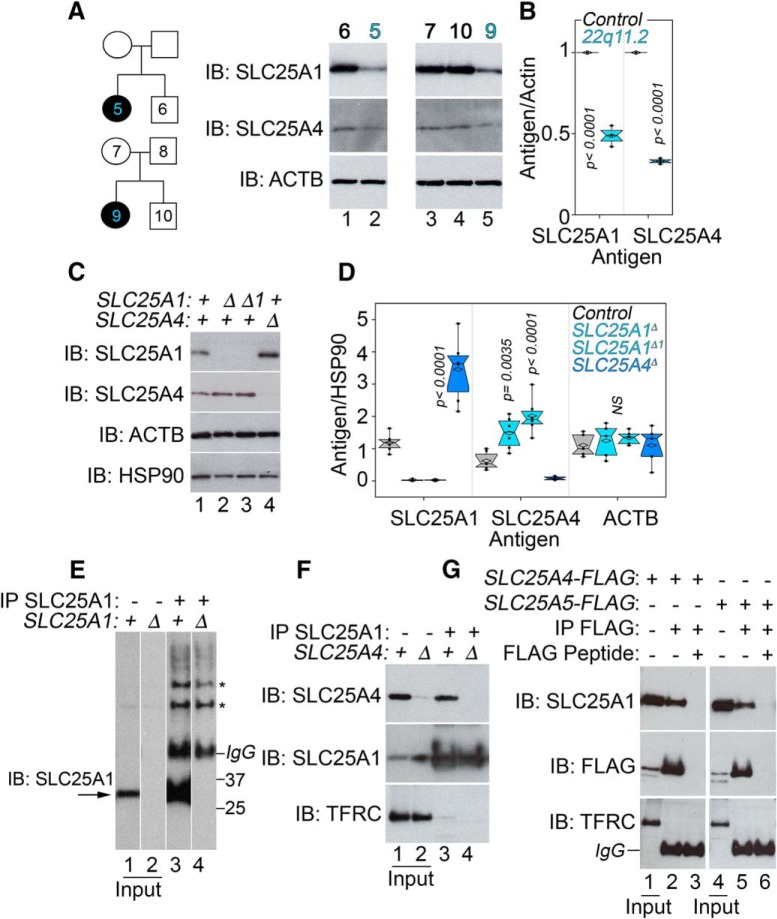
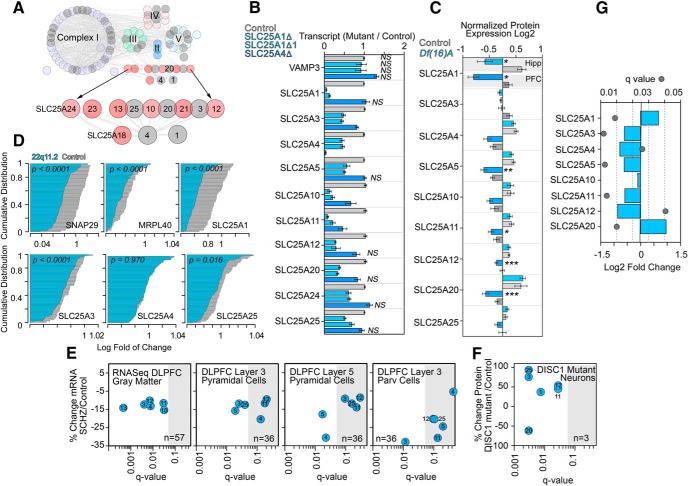
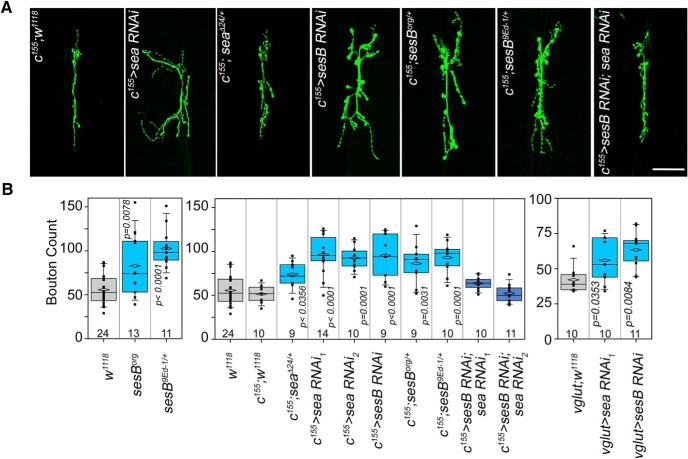
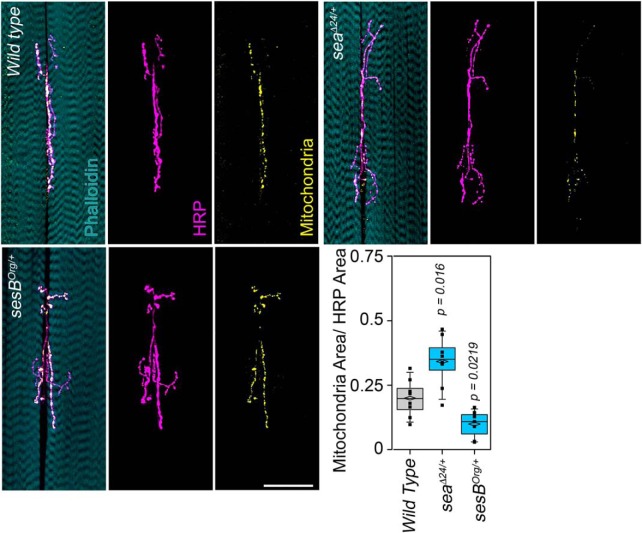
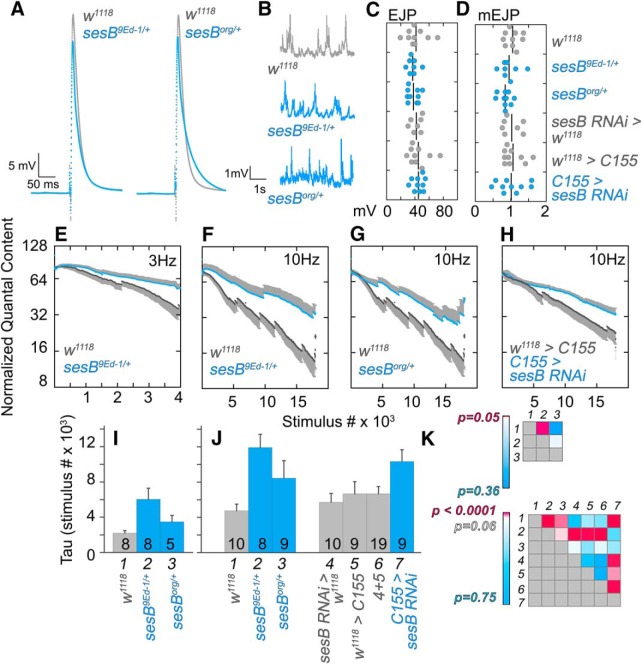
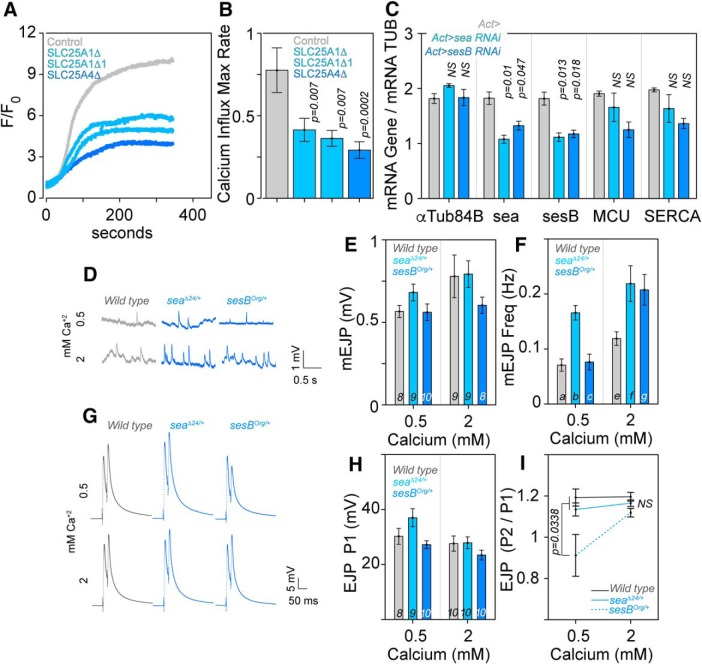
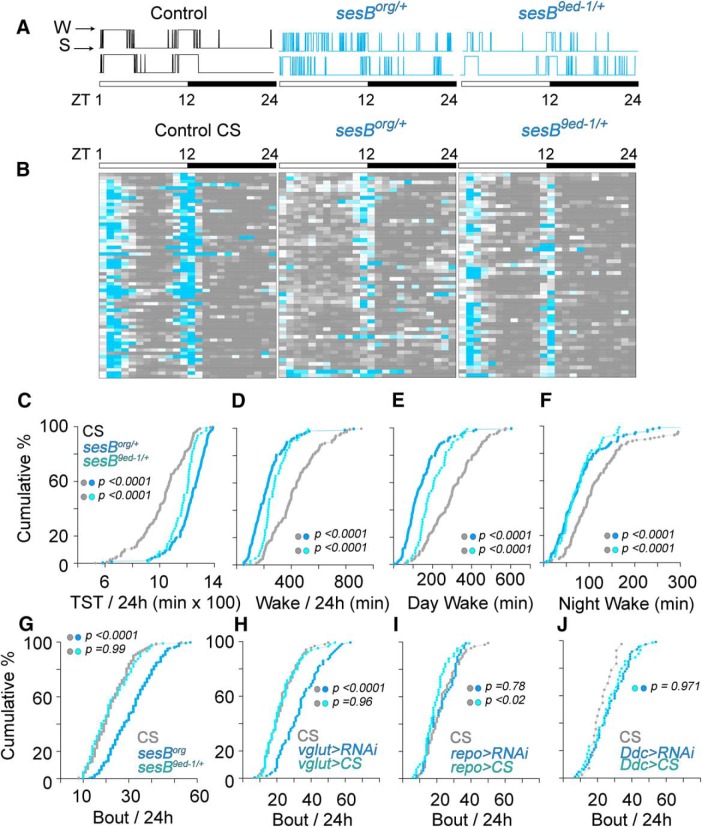
Neurodevelopmental disorders offer insight into synaptic mechanisms. To unbiasedly uncover these mechanisms, we studied the 22q11.2 syndrome, a recurrent copy number variant, which is the highest schizophrenia genetic risk factor. We quantified the proteomes of 22q11.2 mutant human fibroblasts from both sexes and mouse brains carrying a 22q11.2-like defect, Df(16)A+/- Molecular ontologies defined mitochondrial compartments and pathways as some of top ranked categories. In particular, we identified perturbations in the SLC25A1-SLC25A4 mitochondrial transporter interactome as associated with the 22q11.2 genetic defect. Expression of SLC25A1-SLC25A4 interactome components was affected in neuronal cells from schizophrenia patients. Furthermore, hemideficiency of the Drosophila SLC25A1 or SLC25A4 orthologues, dSLC25A1-sea and dSLC25A4-sesB, affected synapse morphology, neurotransmission, plasticity, and sleep patterns. Our findings indicate that synapses are sensitive to partial loss of function of mitochondrial solute transporters. We propose that mitoproteomes regulate synapse development and function in normal and pathological conditions in a cell-specific manner.SIGNIFICANCE STATEMENT We address the central question of how to comprehensively define molecular mechanisms of the most prevalent and penetrant microdeletion associated with neurodevelopmental disorders, the 22q11.2 microdeletion syndrome. This complex mutation reduces gene dosage of ∼63 genes in humans. We describe a disruption of the mitoproteome in 22q11.2 patients and brains of a 22q11.2 mouse model. In particular, we identify a network of inner mitochondrial membrane transporters as a hub required for synapse function. Our findings suggest that mitochondrial composition and function modulate the risk of neurodevelopmental disorders, such as schizophrenia.
Keywords: 22q11.2 microdeletion; SLC25A1; SLC25A4; mitochondria; schizophrenia; synapse.
Copyright © 2019 the authors.
Figures

Similar articles
-
Mitochondrial Proteostasis Requires Genes Encoded in a Neurodevelopmental Syndrome Locus.J Neurosci. 2021 Aug 4;41(31):6596-6616. doi: 10.1523/JNEUROSCI.2197-20.2021. Epub 2021 Jul 14. J Neurosci. 2021. PMID: 34261699 Free PMC article.
-
Mitochondrial Citrate Transporter-dependent Metabolic Signature in the 22q11.2 Deletion Syndrome.J Biol Chem. 2015 Sep 18;290(38):23240-53. doi: 10.1074/jbc.M115.672360. Epub 2015 Jul 28. J Biol Chem. 2015. PMID: 26221035 Free PMC article.
-
Mitochondrial localization and function of a subset of 22q11 deletion syndrome candidate genes.Mol Cell Neurosci. 2008 Nov;39(3):439-51. doi: 10.1016/j.mcn.2008.07.027. Epub 2008 Aug 15. Mol Cell Neurosci. 2008. PMID: 18775783 Free PMC article.
-
Synaptic Plasticity Dysfunctions in the Pathophysiology of 22q11 Deletion Syndrome: Is There a Role for Astrocytes?Int J Mol Sci. 2022 Apr 16;23(8):4412. doi: 10.3390/ijms23084412. Int J Mol Sci. 2022. PMID: 35457231 Free PMC article. Review.
-
Mitochondria in complex psychiatric disorders: Lessons from mouse models of 22q11.2 deletion syndrome: Hemizygous deletion of several mitochondrial genes in the 22q11.2 genomic region can lead to symptoms associated with neuropsychiatric disease.Bioessays. 2017 Feb;39(2):10.1002/bies.201600177. doi: 10.1002/bies.201600177. Epub 2017 Jan 3. Bioessays. 2017. PMID: 28044359 Free PMC article. Review.
Cited by
-
17q12 deletion syndrome mouse model shows defects in craniofacial, brain and kidney development, and glucose homeostasis.Dis Model Mech. 2022 Dec 1;15(12):dmm049752. doi: 10.1242/dmm.049752. Epub 2022 Dec 13. Dis Model Mech. 2022. PMID: 36373506 Free PMC article.
-
The Role of Alpha-Synuclein and Other Parkinson's Genes in Neurodevelopmental and Neurodegenerative Disorders.Int J Mol Sci. 2020 Aug 10;21(16):5724. doi: 10.3390/ijms21165724. Int J Mol Sci. 2020. PMID: 32785033 Free PMC article. Review.
-
FMRP attenuates activity dependent modifications in the mitochondrial proteome.Mol Brain. 2021 Apr 30;14(1):75. doi: 10.1186/s13041-021-00783-w. Mol Brain. 2021. PMID: 33931071 Free PMC article.
-
Review: Genes Involved in Mitochondrial Physiology Within 22q11.2 Deleted Region and Their Relevance to Schizophrenia.Schizophr Bull. 2023 Nov 29;49(6):1637-1653. doi: 10.1093/schbul/sbad066. Schizophr Bull. 2023. PMID: 37379469 Free PMC article.
-
Abnormalities of synaptic mitochondria in autism spectrum disorder and related neurodevelopmental disorders.J Mol Med (Berl). 2021 Feb;99(2):161-178. doi: 10.1007/s00109-020-02018-2. Epub 2020 Dec 18. J Mol Med (Berl). 2021. PMID: 33340060 Free PMC article. Review.
References
-
- Andrews S. (2010) FASTQC: a quality control tool for high throughput sequence data. Available at: http://www.bioinformatics.babraham.ac.uk/projects/fastqc.
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous