

p300/CBP-associated factor promotes autophagic degradation of δ-catenin through acetylation and decreases prostate cancer tumorigenicity
- PMID: 30833716
- PMCID: PMC6399259
- DOI: 10.1038/s41598-019-40238-w
p300/CBP-associated factor promotes autophagic degradation of δ-catenin through acetylation and decreases prostate cancer tumorigenicity
Abstract
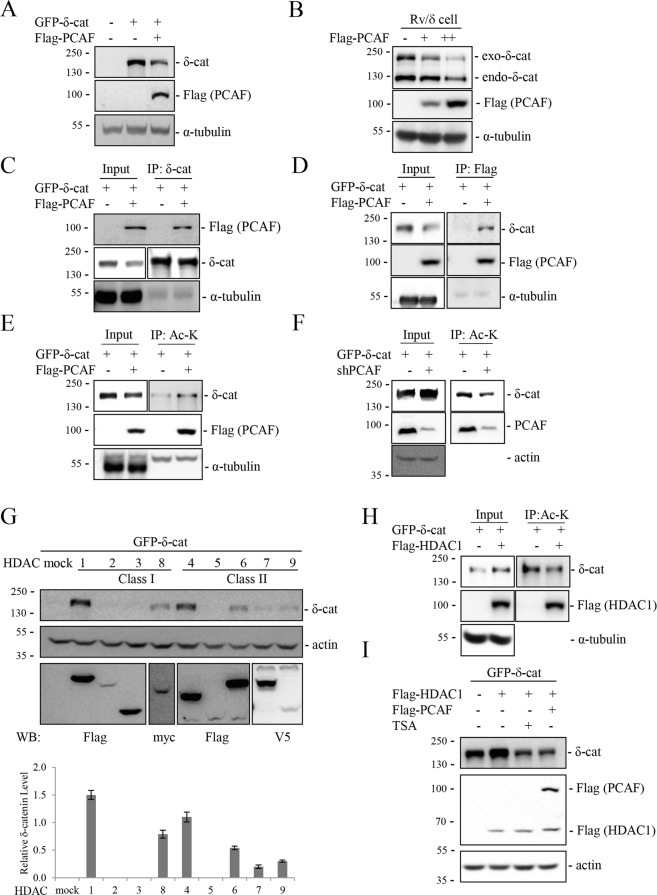
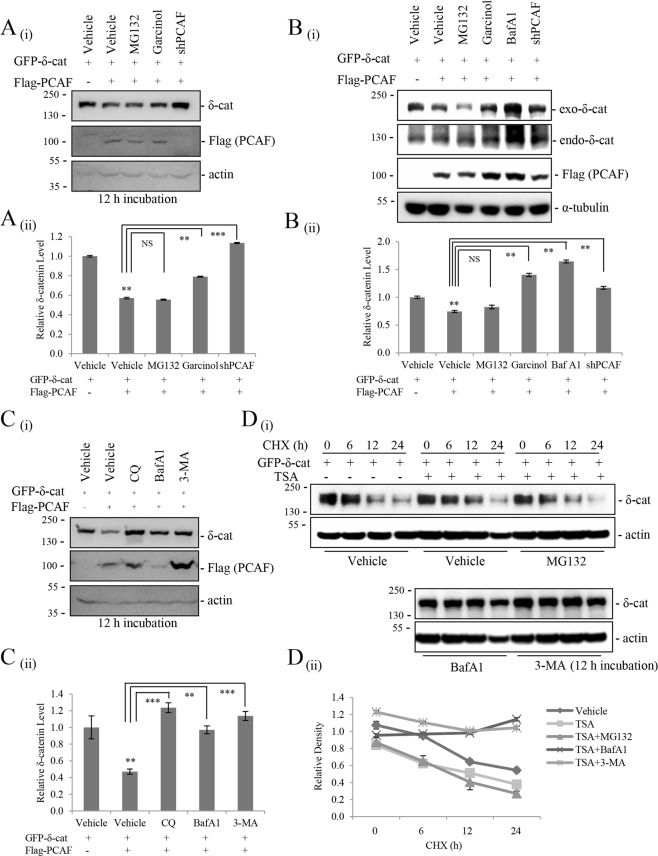
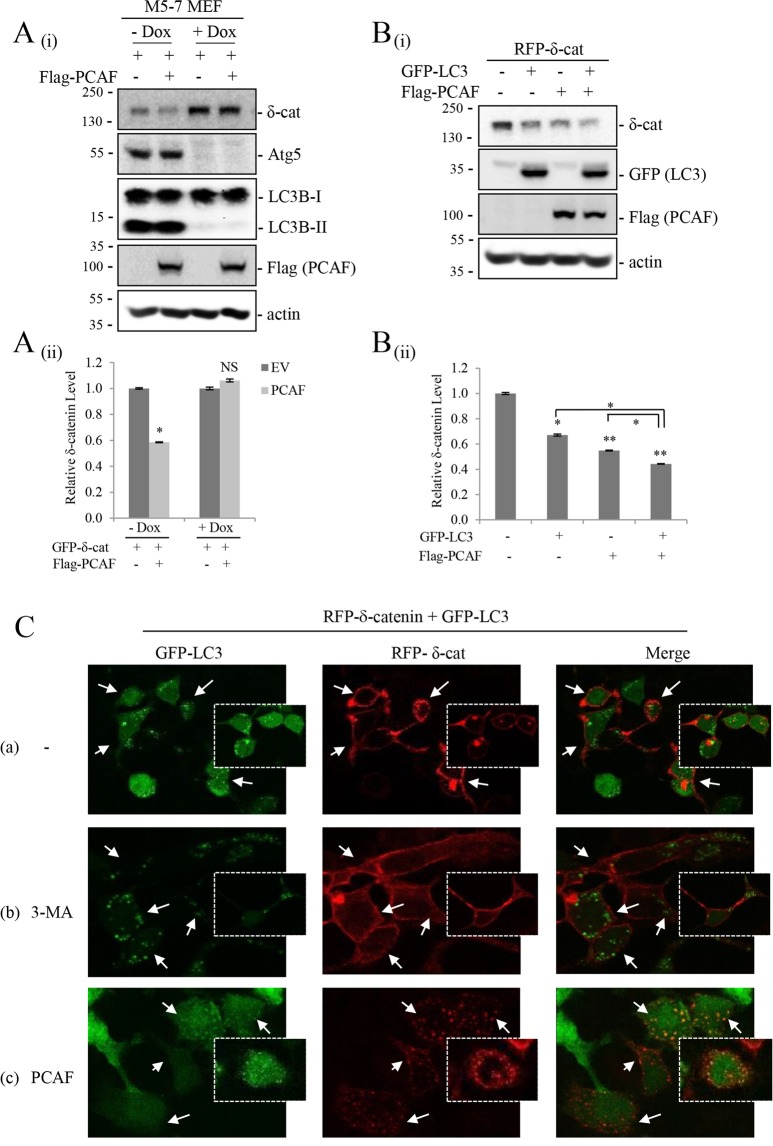
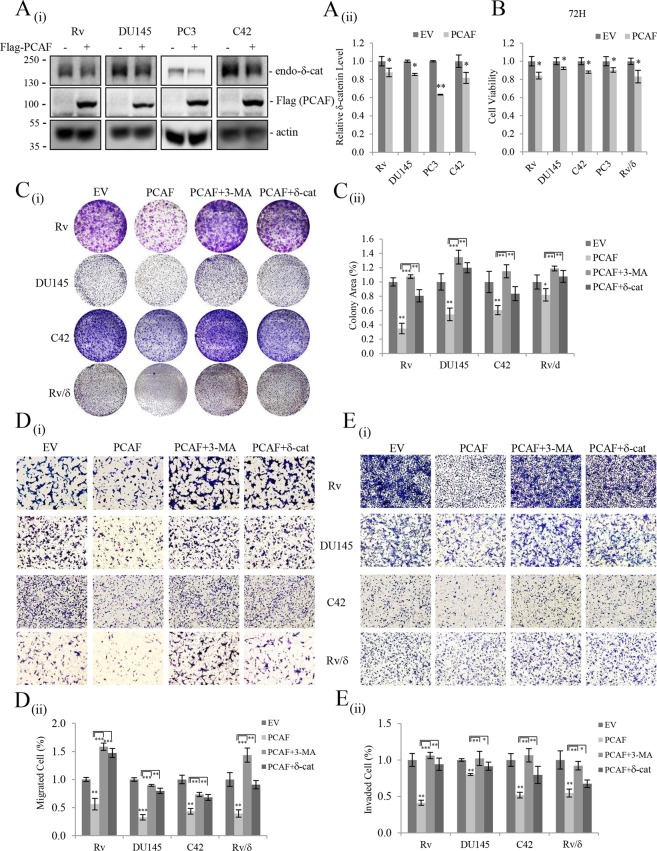
δ-Catenin shares common binding partners with β-catenin. As acetylation and deacetylation regulate β-catenin stability, we searched for histone acetyltransferases (HATs) or histone deacetylases (HDACs) affecting δ-catenin acetylation status and protein levels. We showed that p300/CBP-associated factor (PCAF) directly bound to and acetylated δ-catenin, whereas several class I and class II HDACs reversed this effect. Unlike β-catenin, δ-catenin was downregulated by PCAF-mediated acetylation and upregulated by HDAC-mediated deacetylation. The HDAC inhibitor trichostatin A attenuated HDAC1-mediated δ-catenin upregulation, whereas HAT or autophagy inhibitors, but not proteasome inhibitors, abolished PCAF-mediated δ-catenin downregulation. The results suggested that PCAF-mediated δ-catenin acetylation promotes its autophagic degradation in an Atg5/LC3-dependent manner. Deletions or point mutations identified several lysine residues in different δ-catenin domains involved in PCAF-mediated δ-catenin downregulation. PCAF overexpression in prostate cancer cells markedly reduced δ-catenin levels and suppressed cell growth and motility. PCAF-mediated δ-catenin downregulation inhibited E-cadherin processing and decreased the nuclear distribution of β-catenin, resulting in the suppression of β-catenin/LEF-1-mediated downstream effectors. These data demonstrate that PCAF downregulates δ-catenin by promoting its autophagic degradation and suppresses δ-catenin-mediated oncogenic signals.
Conflict of interest statement
The authors declare no competing interests.
Figures



Similar articles
-
P300/CBP-associated factor (PCAF)-mediated acetylation of Fascin at lysine 471 inhibits its actin-bundling activity and tumor metastasis in esophageal cancer.Cancer Commun (Lond). 2021 Dec;41(12):1398-1416. doi: 10.1002/cac2.12221. Epub 2021 Sep 23. Cancer Commun (Lond). 2021. PMID: 34555274 Free PMC article.
-
Wnt/β-catenin-dependent acetylation of Pygo2 by CBP/p300 histone acetyltransferase family members.Biochem J. 2016 Nov 15;473(22):4193-4203. doi: 10.1042/BCJ20160590. Epub 2016 Sep 19. Biochem J. 2016. PMID: 27647933
-
Trichostatin A induces transforming growth factor beta type II receptor promoter activity and acetylation of Sp1 by recruitment of PCAF/p300 to a Sp1.NF-Y complex.J Biol Chem. 2005 Mar 18;280(11):10047-54. doi: 10.1074/jbc.M408680200. Epub 2005 Jan 12. J Biol Chem. 2005. PMID: 15647279
-
Regulation of protein turnover by acetyltransferases and deacetylases.Biochimie. 2008 Feb;90(2):306-12. doi: 10.1016/j.biochi.2007.06.009. Epub 2007 Jul 1. Biochimie. 2008. PMID: 17681659 Review.
-
The functional significance of nuclear receptor acetylation.Steroids. 2007 Feb;72(2):221-30. doi: 10.1016/j.steroids.2006.12.001. Epub 2007 Feb 7. Steroids. 2007. PMID: 17291555 Free PMC article. Review.
Cited by
-
P300 promotes tumor recurrence by regulating radiation-induced conversion of glioma stem cells to vascular-like cells.Nat Commun. 2022 Oct 19;13(1):6202. doi: 10.1038/s41467-022-33943-0. Nat Commun. 2022. PMID: 36261421 Free PMC article.
-
Targeting autophagy in prostate cancer: preclinical and clinical evidence for therapeutic response.J Exp Clin Cancer Res. 2022 Mar 22;41(1):105. doi: 10.1186/s13046-022-02293-6. J Exp Clin Cancer Res. 2022. PMID: 35317831 Free PMC article. Review.
-
KITENIN promotes aerobic glycolysis through PKM2 induction by upregulating the c-Myc/hnRNPs axis in colorectal cancer.Cell Biosci. 2023 Aug 8;13(1):146. doi: 10.1186/s13578-023-01089-1. Cell Biosci. 2023. PMID: 37553596 Free PMC article.
-
Delta-Catenin as a Modulator of Rho GTPases in Neurons.Front Cell Neurosci. 2022 Jul 4;16:939143. doi: 10.3389/fncel.2022.939143. eCollection 2022. Front Cell Neurosci. 2022. PMID: 35860313 Free PMC article. Review.
-
Bacterial Lipopolysaccharide Augmented Malignant Transformation and Promoted the Stemness in Prostate Cancer Epithelial Cells.J Inflamm Res. 2021 Nov 9;14:5849-5862. doi: 10.2147/JIR.S332943. eCollection 2021. J Inflamm Res. 2021. PMID: 34785925 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
