

RAD51D splice variants and cancer-associated mutations reveal XRCC2 interaction to be critical for homologous recombination
- PMID: 30836272
- PMCID: PMC6508892
- DOI: 10.1016/j.dnarep.2019.02.008
RAD51D splice variants and cancer-associated mutations reveal XRCC2 interaction to be critical for homologous recombination
Abstract
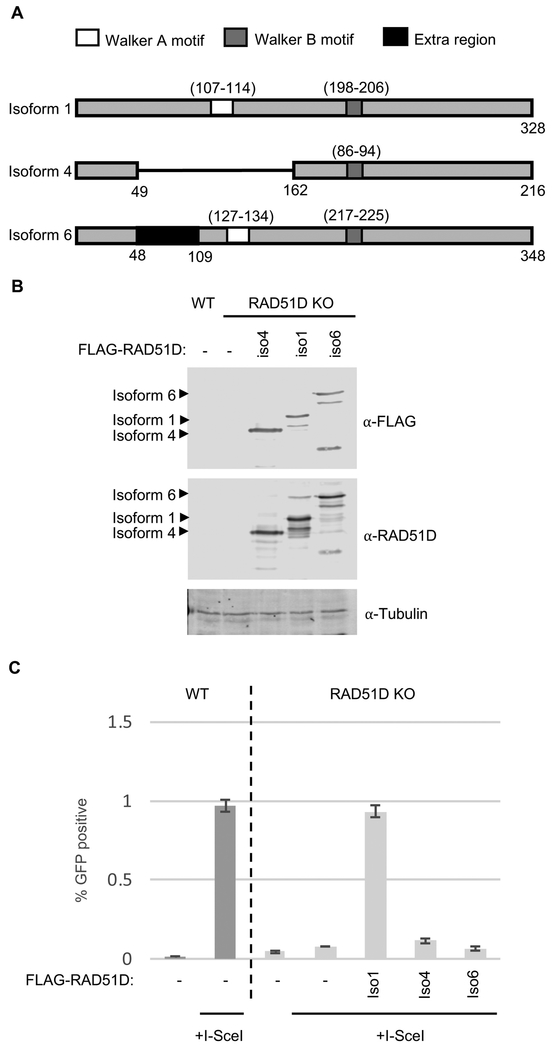
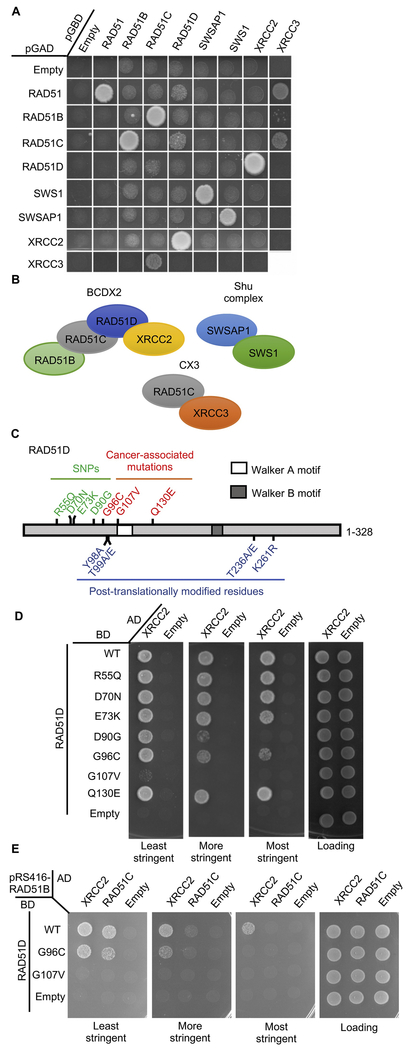
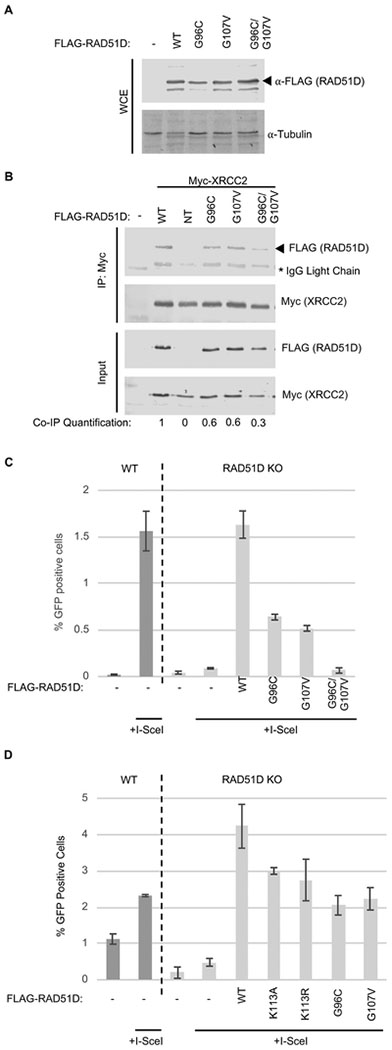
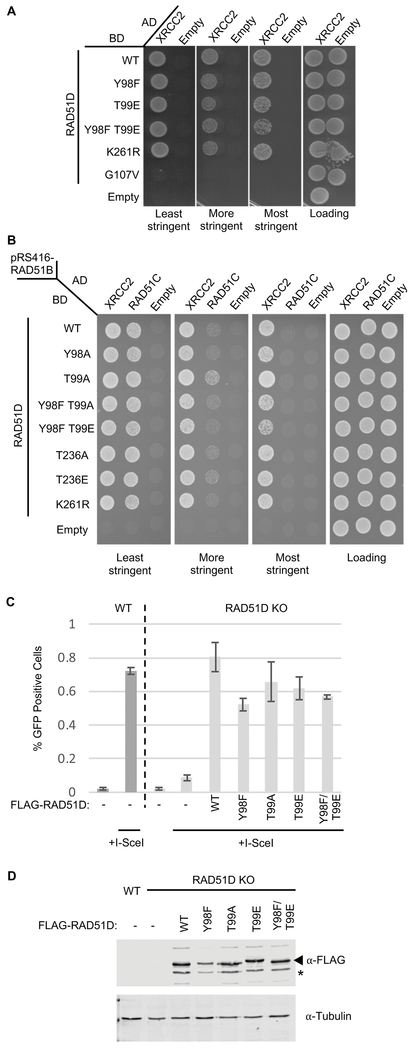
The proficiency of cancer cells to repair DNA double-strand breaks (DSBs) by homologous recombination (HR) is a key determinant in predicting response to targeted therapies such as PARP inhibitors. The RAD51 paralogs work as multimeric complexes and act downstream of BRCA1 to facilitate HR. Numerous epidemiological studies have linked RAD51 paralog mutations with hereditary cancer predisposition. Despite their substantial links to cancer, RAD51 paralog HR function has remained elusive. Here we identify isoform 1 as the functional isoform of RAD51D, whereas isoform 4 which has a large N-terminal deletion (including the Walker A motif), and isoform 6 which includes an alternate exon in the N-terminus, are non-functional. To determine the importance of this N-terminal region, we investigated the impact of cancer-associated mutations and SNPs in this variable RAD51D N-terminal region using yeast-2-hybrid and yeast-3-hybrid assays to screen for altered protein-protein interactions. We identified two cancer-associated mutations close to or within the Walker A motif (G96C and G107 V, respectively) that independently disrupt RAD51D interaction with XRCC2. We validated our yeast interaction data in human U2OS cells by co-immunoprecipitation and determined the impact of these mutations on HR-proficiency using a sister chromatid recombination reporter assay in a RAD51D knock-out cell line. Our investigation reveals that the interaction of RAD51D with XRCC2 is required for DSB repair. By characterizing the impact of cancer-associated mutations on RAD51D interactions, we aim to develop predictive models for therapeutic sensitivity and resistance in patients who harbor similar mutations in RAD51D.
Keywords: Double-strand break repair; Homologous recombination; RAD51 paralogs; RAD51D; Walker A motif; XRCC2.
Copyright © 2019 Elsevier B.V. All rights reserved.
Conflict of interest statement
Conflicts of interest
The authors declare that there are no conflicts of interest.
Figures
References
-
- Renkawitz J, Lademann CA, and Jentsch S, Mechanisms and principles of homology search during recombination. Nat Rev Mol Cell Biol, 2014. 15(6): p. 369–383. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
