

Nucleation and Formation of a Primary Clot in Insect Blood
- PMID: 30837584
- PMCID: PMC6401176
- DOI: 10.1038/s41598-019-40129-0
Nucleation and Formation of a Primary Clot in Insect Blood
Abstract
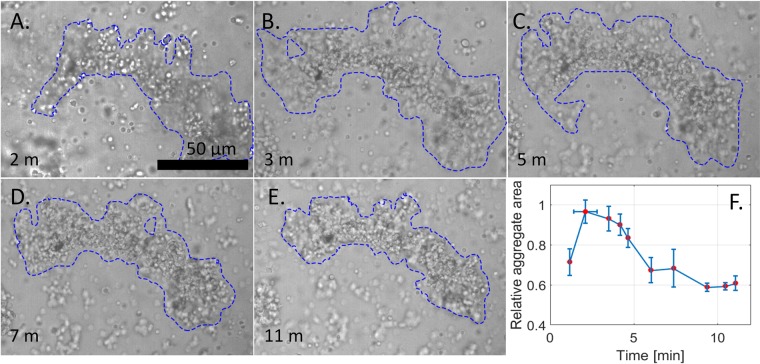
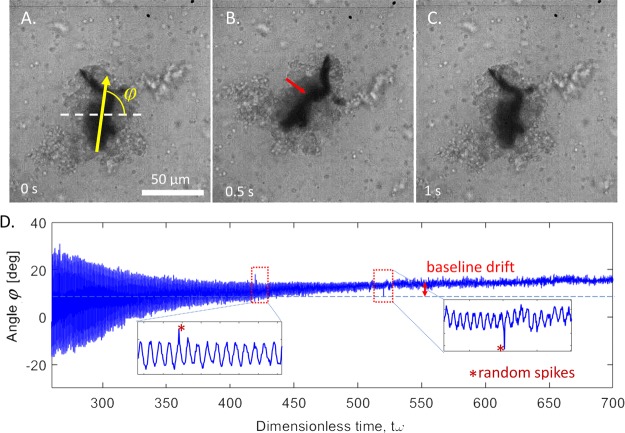
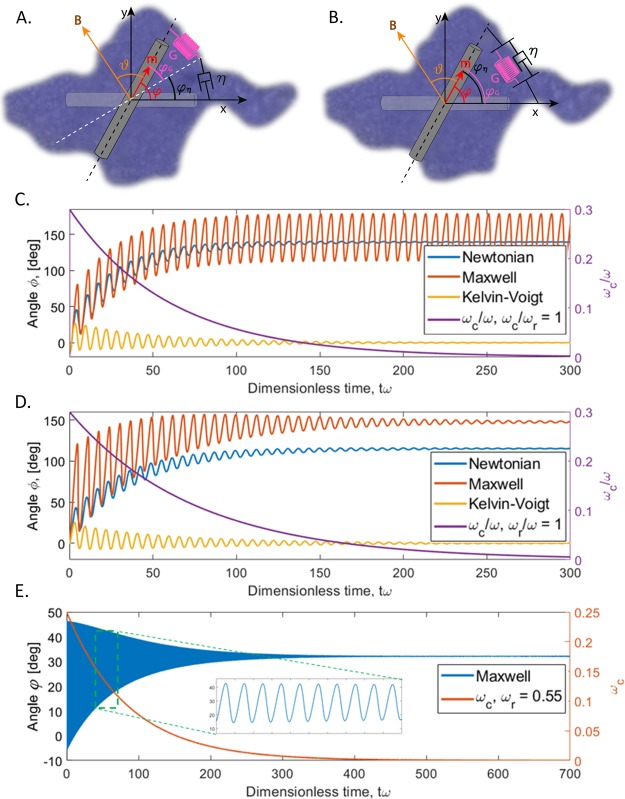
Blood clotting at wound sites is critical for preventing blood loss and invasion by microorganisms in multicellular animals, especially small insects vulnerable to dehydration. The mechanistic reaction of the clot is the first step in providing scaffolding for the formation of new epithelial and cuticular tissue. The clot, therefore, requires special materials properties. We have developed and used nano-rheological magnetic rotational spectroscopy with nanorods to quantitatively study nucleation of cell aggregates that occurs within fractions of a second. Using larvae of Manduca sexta, we discovered that clot nucleation is a two-step process whereby cell aggregation is the time-limiting step followed by rigidification of the aggregate. Clot nucleation and transformation of viscous blood into a visco-elastic aggregate happens in a few minutes, which is hundreds of times faster than wound plugging and scab formation. This discovery sets a time scale for insect clotting phenomena, establishing a materials metric for the kinetics of biochemical reaction cascades. Combined with biochemical and biomolecular studies, these discoveries can help design fast-working thickeners for vertebrate blood, including human blood, based on clotting principles of insect blood.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Salt, G. The cellular defence reactions of insects. (Cambridge University Press, 1970).
-
- Jiang, H., Vilcinskas, A. & Kanost, M. R. In Invertebrate Immunity Vol. 708 Advances in experimental medicine and biology (ed K. Soderhall) 181–204 (Springer-Verlag Berlin, 2010).
-
- Strand MR. The insect cellular immune response. Insect Sci. 2008;15:1–14. doi: 10.1111/j.1744-7917.2008.00183.x. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
