

Identifying and Engineering Genes for Parthenogenesis in Plants
- PMID: 30838007
- PMCID: PMC6389702
- DOI: 10.3389/fpls.2019.00128
Identifying and Engineering Genes for Parthenogenesis in Plants
Abstract
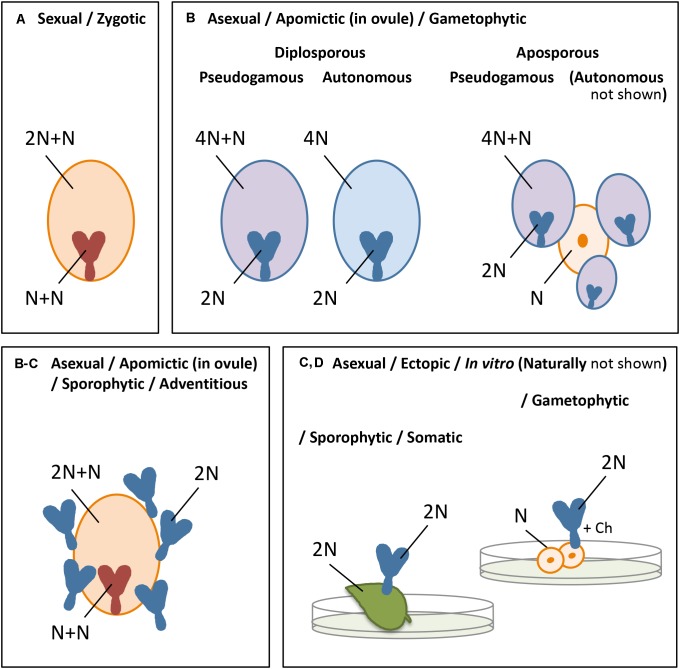
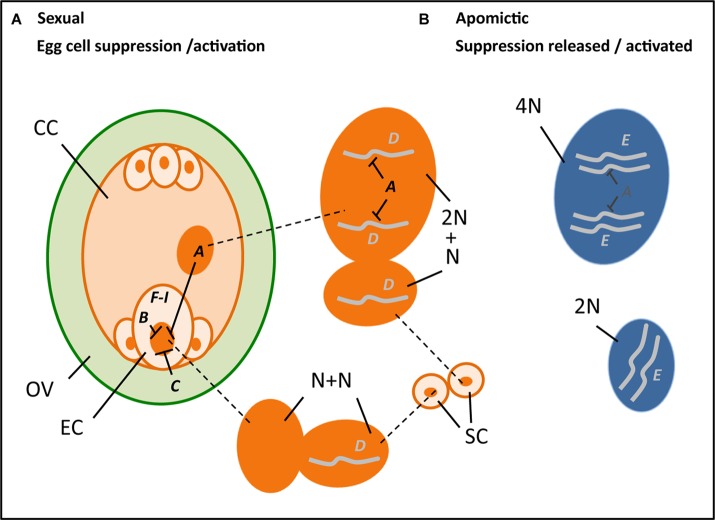
Parthenogenesis is the spontaneous development of an embryo from an unfertilized egg cell. It naturally occurs in a variety of plant and animal species. In plants, parthenogenesis usually is found in combination with apomeiosis (the omission of meiosis) and pseudogamous or autonomous (with or without central cell fertilization) endosperm formation, together known as apomixis (clonal seed production). The initiation of embryogenesis in vivo and in vitro has high potential in plant breeding methods, particularly for the instant production of homozygous lines from haploid gametes [doubled haploids (DHs)], the maintenance of vigorous F1-hybrids through clonal seed production after combining it with apomeiosis, reverse breeding approaches, and for linking diploid and polyploid gene pools. Because of this large interest, efforts to identify gene(s) for parthenogenesis from natural apomicts have been undertaken by using map-based cloning strategies and comparative gene expression studies. In addition, engineering parthenogenesis in sexual model species has been investigated via mutagenesis and gain-of-function strategies. These efforts have started to pay off, particularly by the isolation of the PsASGR-BabyBoom-Like from apomictic Pennisetum, a gene proven to be transferable to and functional in sexual pearl millet, rice, and maize. This review aims to summarize the current knowledge on parthenogenesis, the possible gene candidates also outside the grasses, and the use of these genes in plant breeding protocols. It shows that parthenogenesis is able to inherit and function independently from apomeiosis and endosperm formation, is expressed and active in the egg cell, and can induce embryogenesis in polyploid, diploid as well as haploid egg cells in plants. It also shows the importance of genes involved in the suppression of transcription and modifications thereof at one hand, and in embryogenesis for which transcription is allowed or artificially overexpressed on the other, in parthenogenetic reproduction. Finally, it emphasizes the importance of functional endosperm to allow for successful embryo growth and viable seed production.
Keywords: Pennisetum; PsASGR-BabyBoom-Like (PsASGR-BBML); Taraxacum; apomixis; doubled haploids; embryo induction; embryogenesis; parthenogenesis.
Figures
References
-
- Agashe B., Prasad C. K., Siddiqi I. (2002). Identification and analysis of DYAD: a gene required for meiotic chromosome organisation and female meiotic progression in Arabidopsis. Development 129 3935–3943. - PubMed
-
- Akiyama Y., Conner J. A., Goel S., Morishige D. T., Mullet J. E., Hanna W. W., et al. (2004). High-resolution physical mapping in Pennisetum squamulatum reveals extensive chromosomal heteromorphism of the genomic region associated with apomixis. Plant Physiol. 134 1733–1741. 10.1104/pp.103.033969 - DOI - PMC - PubMed
-
- Anderson S. N., Johnson C. S., Jones D. S., Conrad L. J., Gou X., Russell S. D., et al. (2013). Transcriptomes of isolated Oryza sativa gametes characterized by deep sequencing: evidence for distinct sex-dependent chromatin and epigenetic states before fertilization. Plant J. 76 729–741. 10.1111/tpj.12336 - DOI - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources