

Epigenetic regulation of REX1 expression and chromatin binding specificity by HMGNs
- PMID: 30838422
- PMCID: PMC6511878
- DOI: 10.1093/nar/gkz161
Epigenetic regulation of REX1 expression and chromatin binding specificity by HMGNs
Abstract
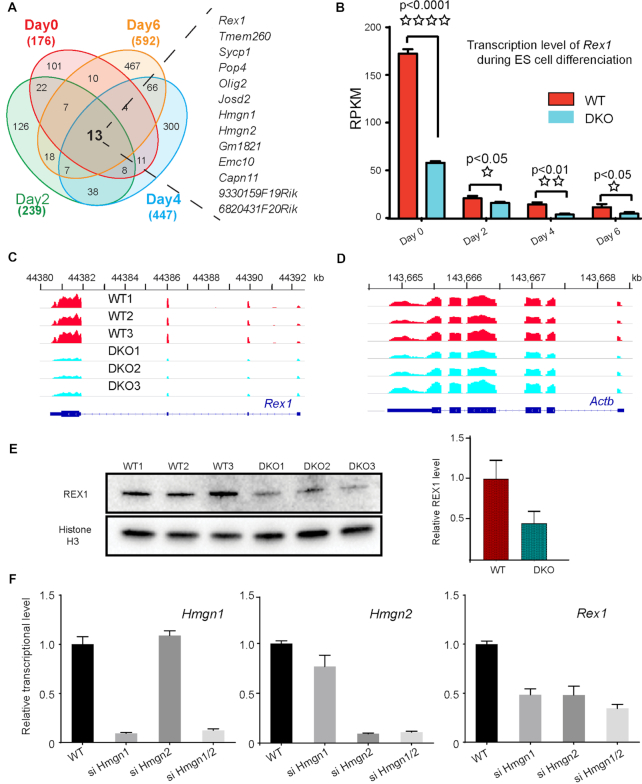
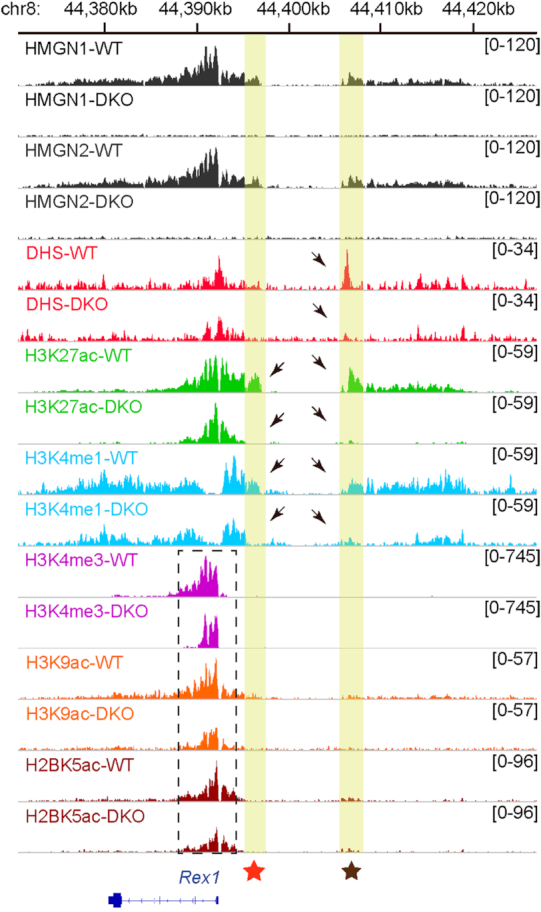
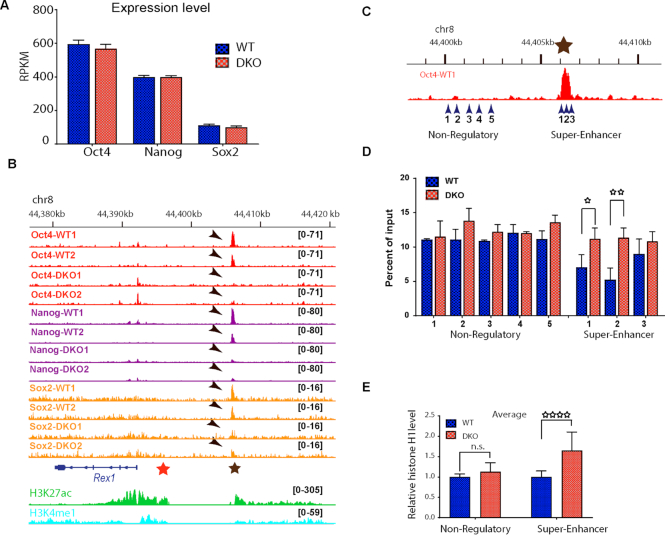
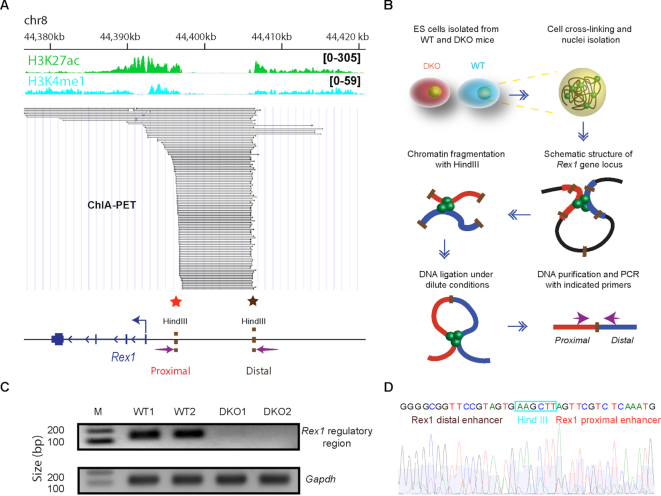
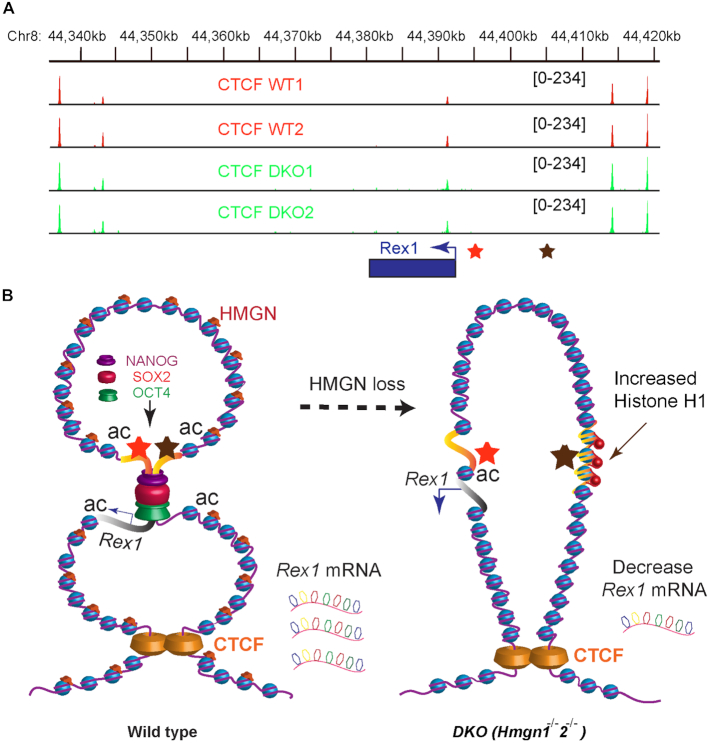
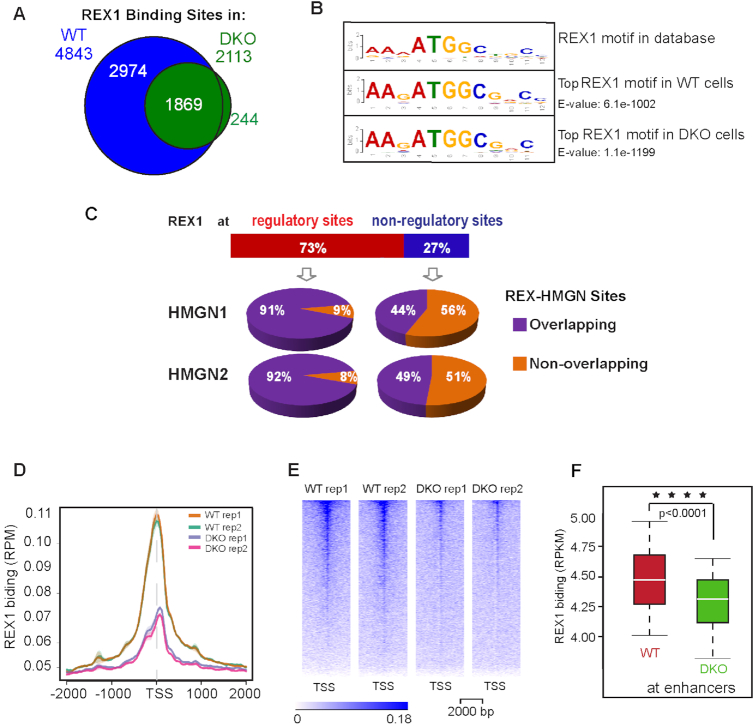
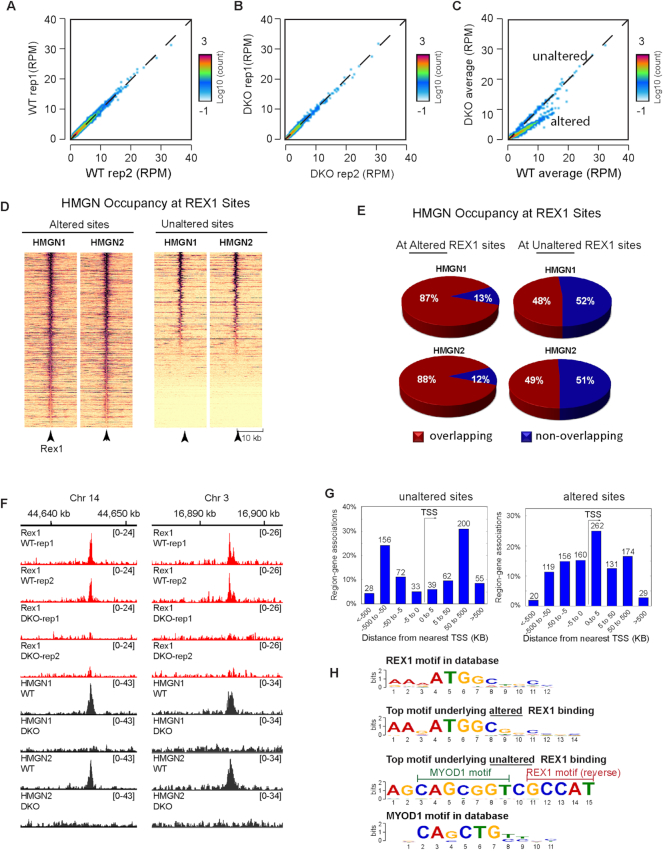
HMGN proteins localize to chromatin regulatory sites and modulate the cell-type specific transcription profile; however, the molecular mechanism whereby these ubiquitous nucleosome binding proteins affect gene expression is not fully understood. Here, we show that HMGNs regulate the expression of Rex1, one of the most highly transcribed genes in mouse embryonic stem cells (ESCs), by recruiting the transcription factors NANOG, OCT4 and SOX2 to an ESC-specific super enhancer located in the 5' region of Rex1. HMGNs facilitate the establishment of an epigenetic landscape characteristic of active chromatin and enhancer promoter interactions, as seen by chromatin conformation capture. Loss of HMGNs alters the local epigenetic profile, increases histone H1 occupancy, decreases transcription factors binding and reduces enhancer promoter interactions, thereby downregulating, but not abolishing Rex1 expression. ChIP-seq analyses show high colocalization of HMGNs and of REX1, a zinc finger protein, at promoters and enhancers. Loss of HMGNs preferentially reduces the specific binding of REX1 to these chromatin regulatory sites. Thus, HMGNs affects both the expression and the chromatin binding specificity of REX1. We suggest that HMGNs affect cell-type specific gene expression by modulating the binding specificity of transcription factors to chromatin.
Published by Oxford University Press on behalf of Nucleic Acids Research 2019.
Figures
References
-
- Izzo A., Schneider R.. The role of linker histone H1 modifications in the regulation of gene expression and chromatin dynamics. Biochim. Biophys. Acta. 2016; 1859:486–495. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
