

Examination of the role of sphingosine kinase 2 in a murine model of systemic lupus erythematosus
- PMID: 30840833
- PMCID: PMC6529345
- DOI: 10.1096/fj.201802535R
Examination of the role of sphingosine kinase 2 in a murine model of systemic lupus erythematosus
Abstract
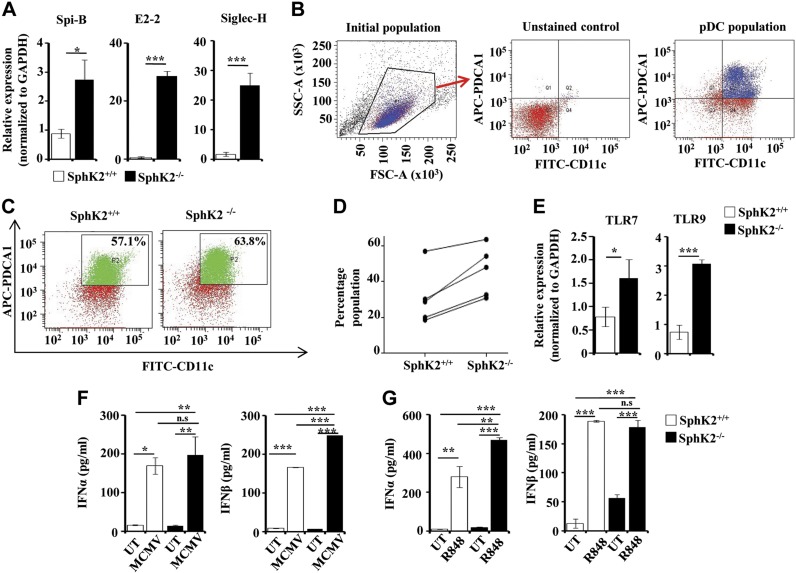
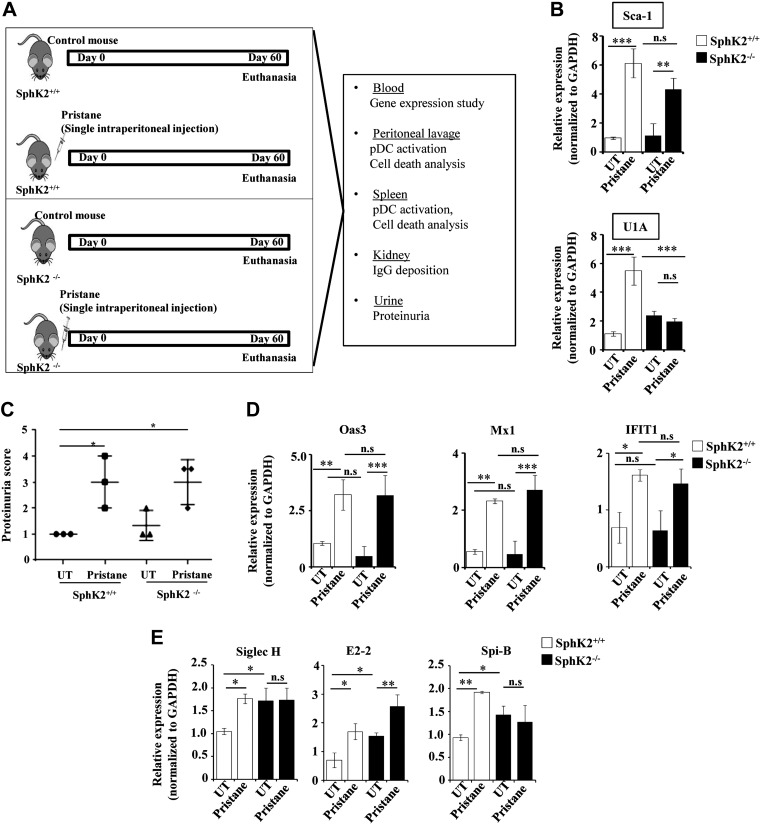
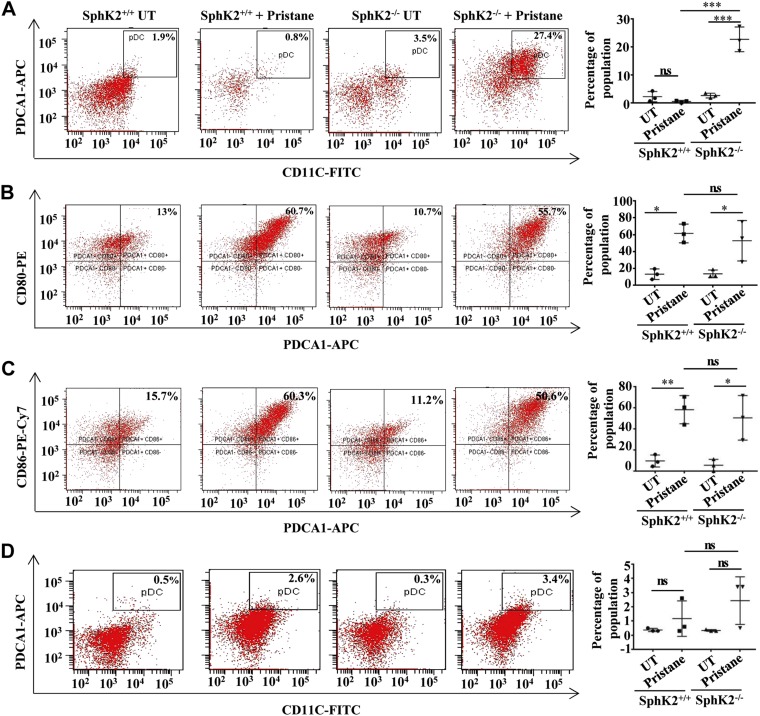
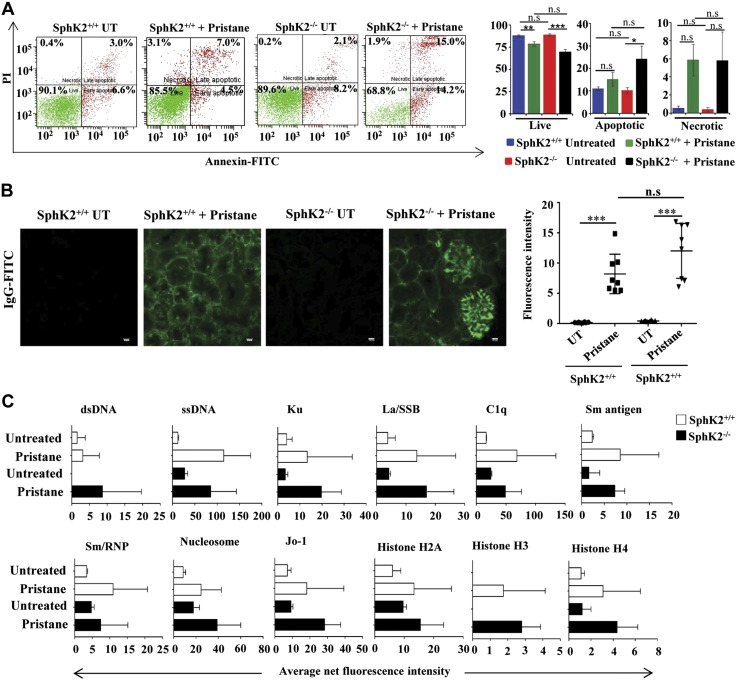
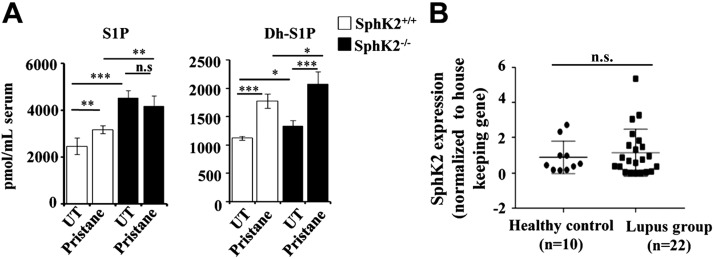
Systemic lupus erythematosus is an autoimmune disease characterized by overproduction of type 1 IFN that causes multiple organ dysfunctions. Plasmacytoid dendritic cells (pDCs) that secrete large amounts of IFN have recently been implicated in the initiation of the disease in preclinical mouse models. Sphingosine-1-phosphate, a bioactive sphingolipid metabolite, is produced by 2 highly conserved isoenzymes, sphingosine kinase (SphK) 1 and SphK2, and regulates diverse processes important for immune responses and autoimmunity. However, not much is known about the role of SphK2 in autoimmune disorders. In this work, we examined the role of SphK2 in pDC development and activation and in the pristane-induced lupus model in mice that mimics the hallmarks of the human disease. Increases in pDC-specific markers were observed in peripheral blood of SphK2 knockout mice. In agreement, the absence of SphK2 increased the differentiation of FMS-like tyrosine kinase 3 ligand dendritic cells as well as expression of endosomal TLRs, TLR7 and TLR9, that modulate production of IFN. Surprisingly, however, SphK2 deficiency did not affect the initiation or progression of pristane-induced lupus. Moreover, although absence of SphK2 increased pDC frequency in pristane-induced lupus, there were no major changes in their activation status. Additionally, SphK2 expression was unaltered in lupus patients. Taken together, our results suggest that SphK2 may play a role in dendritic cell development. Yet, because its deletion had no effect on the clinical lupus parameters in this preclinical model, inhibitors of SphK2 might not be useful for treatment of this devastating disease.-Mohammed, S., Vineetha, N. S., James, S., Aparna, J. S., Lankadasari, M. B., Allegood, J. C., Li, Q.-Z., Spiegel, S., Harikumar, K. B. Examination of the role of sphingosine kinase 2 in a murine model of systemic lupus erythematosus.
Keywords: IFN; S1P; autoimmunity; pDCs; pristane.
Conflict of interest statement
The authors are thankful to the personnel of the Flow Cytometry Core Facility and Bioimaging Facility for excellent technical assistance, and the Animal Research Facility for mouse colony maintenance. S.M. and M.B.L. acknowledge the senior research fellowship from the Indian Council of Medical Research and the University Grant Commission, respectively. S.M. acknowledges the Doctoral Advisory Committee for valuable suggestions. The work was supported by the U.S. National Institutes of Health, National Institute of General Medical Sciences Grant R01GM043880 (to S.S.), a fast-track grant from the Indian Government’s Department of Science and Technology (SR/FT/LS-159/2012), a grand-in-aid scheme of the Council for Scientific and Industrial Research [37 (1720)/18/EMR-II], and, in part, by a faculty start-up grant and Department of Biotechnology (DBT) Ramalingaswami Re-Entry Fellowship (BT/RLF/Re-entry/38/2011) to K.B.H. The authors declare no conflicts of interest.
Figures
References
-
- Siegal F. P., Kadowaki N., Shodell M., Fitzgerald-Bocarsly P. A., Shah K., Ho S., Antonenko S., Liu Y. J. (1999) The nature of the principal type 1 interferon-producing cells in human blood. Science 284, 1835–1837 - PubMed
-
- Rönnblom L., Eloranta M. L., Alm G. V. (2006) The type I interferon system in systemic lupus erythematosus. Arthritis Rheum. 54, 408–420 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
