

Nitroxyl (HNO) targets phospholamban cysteines 41 and 46 to enhance cardiac function
- PMID: 30842219
- PMCID: PMC6571998
- DOI: 10.1085/jgp.201812208
Nitroxyl (HNO) targets phospholamban cysteines 41 and 46 to enhance cardiac function
Abstract
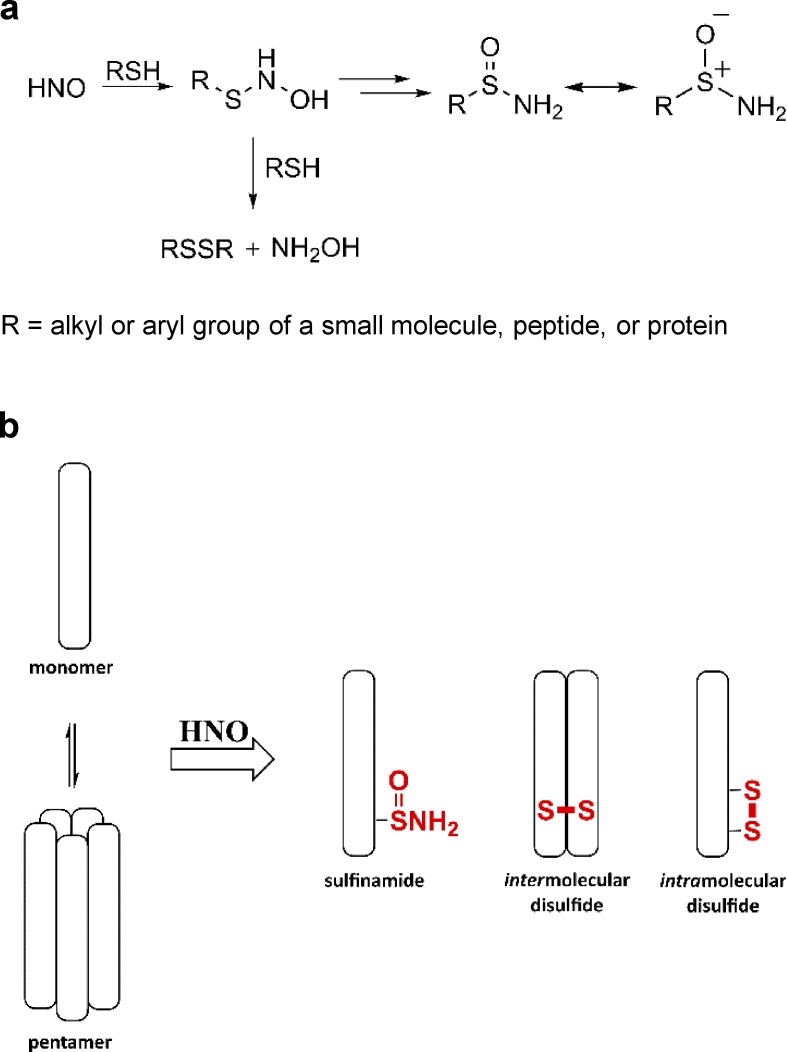
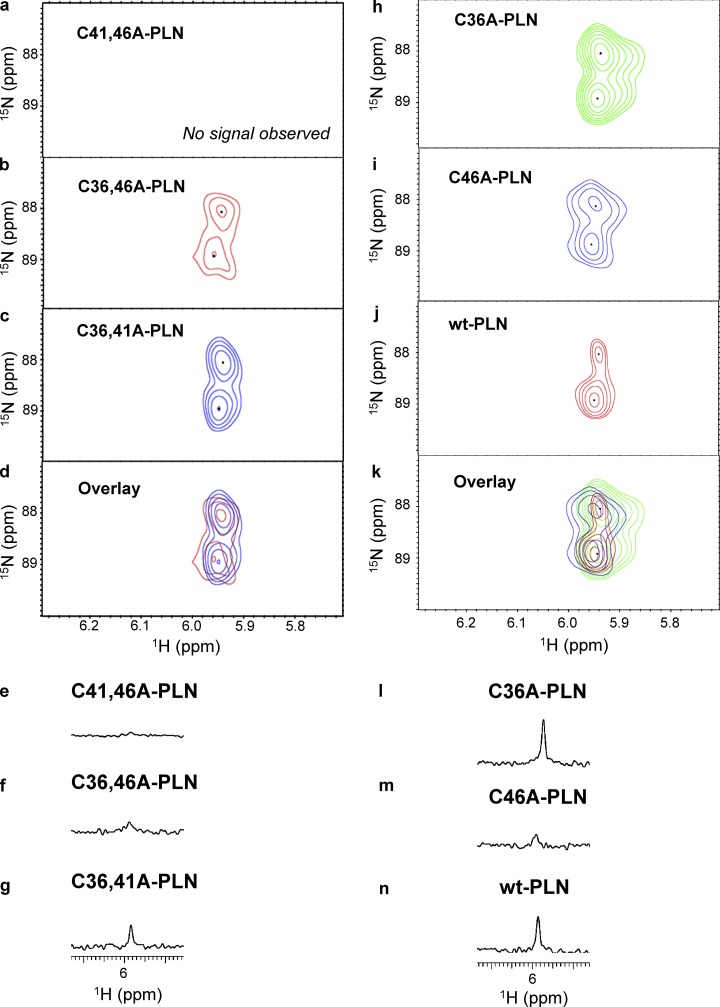
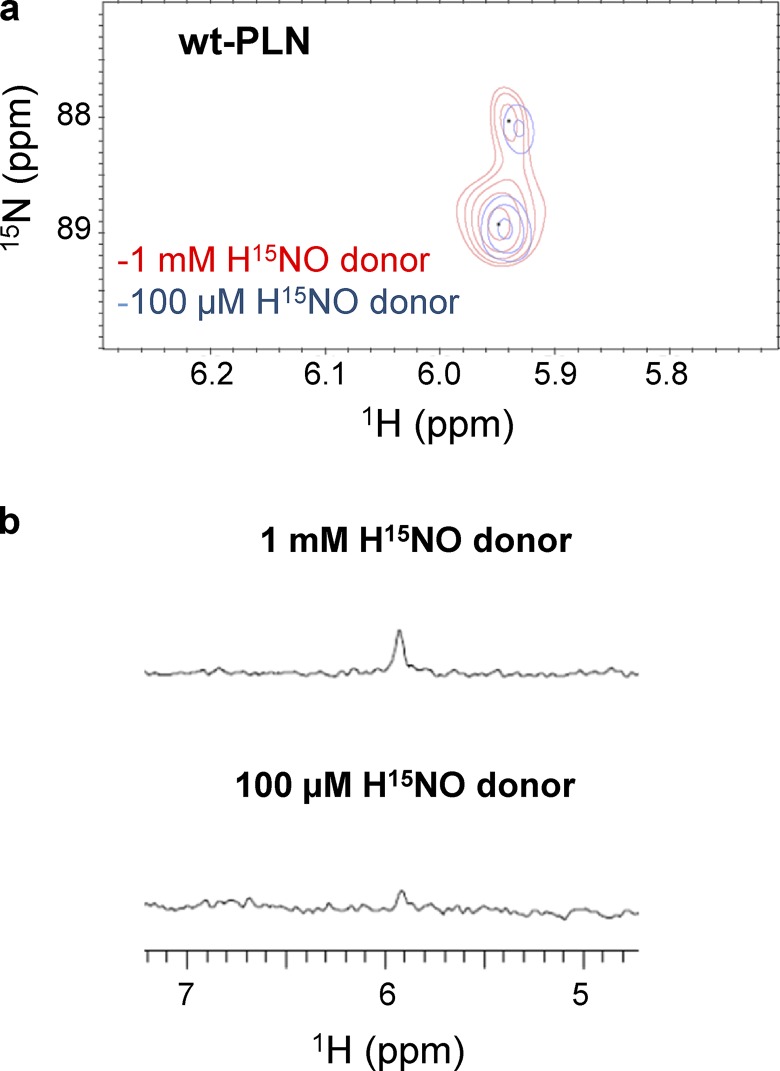
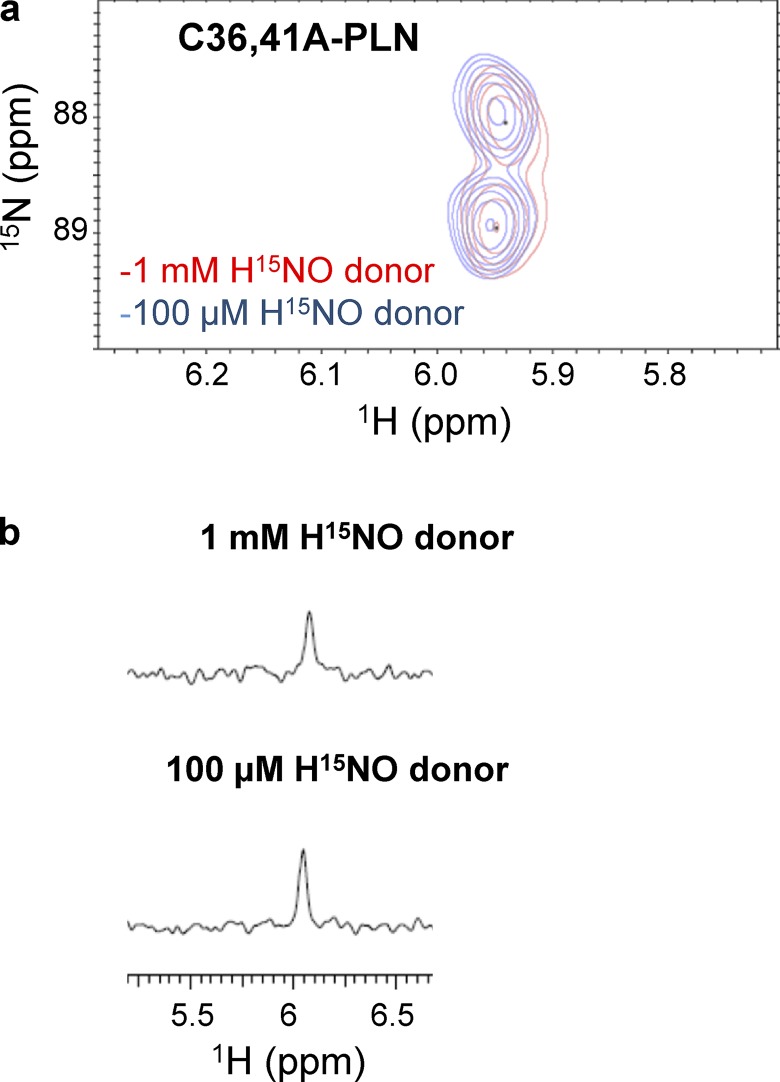
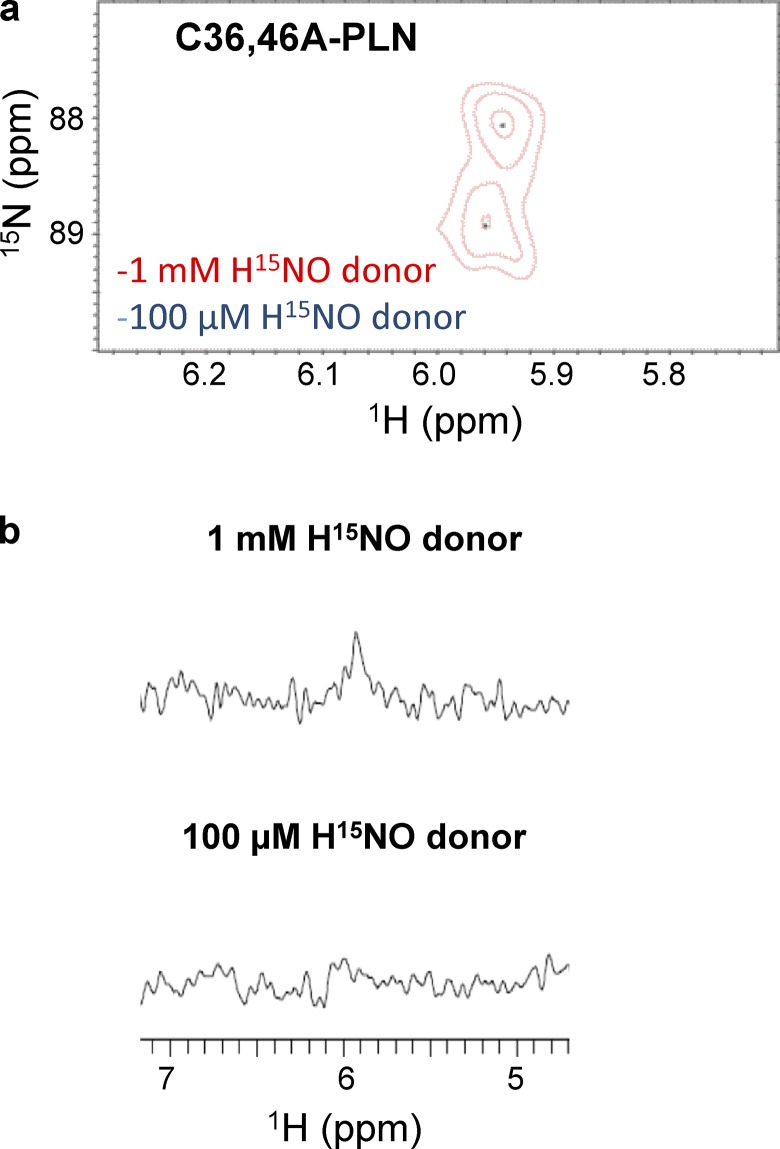
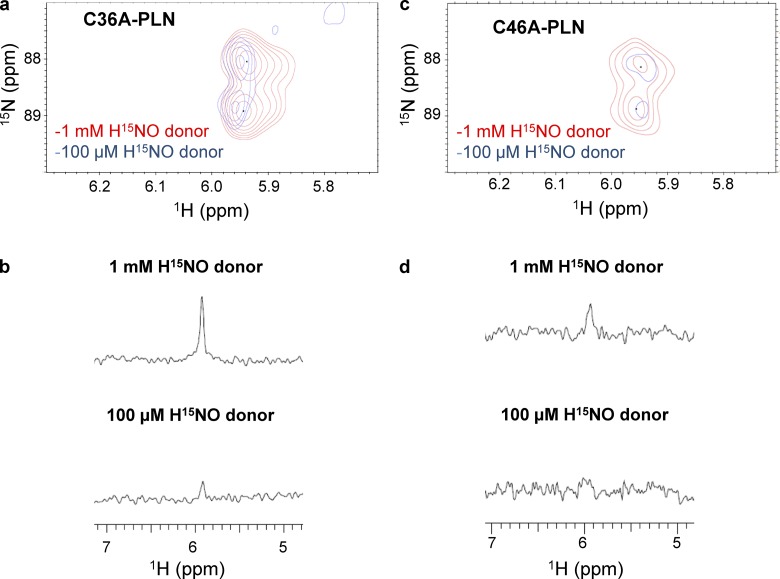
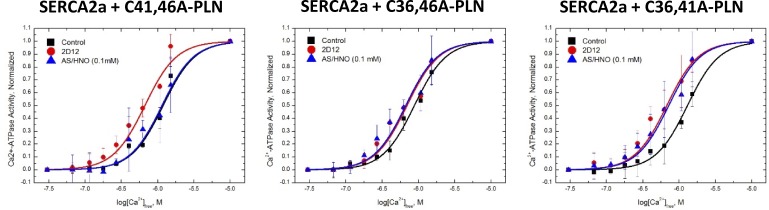
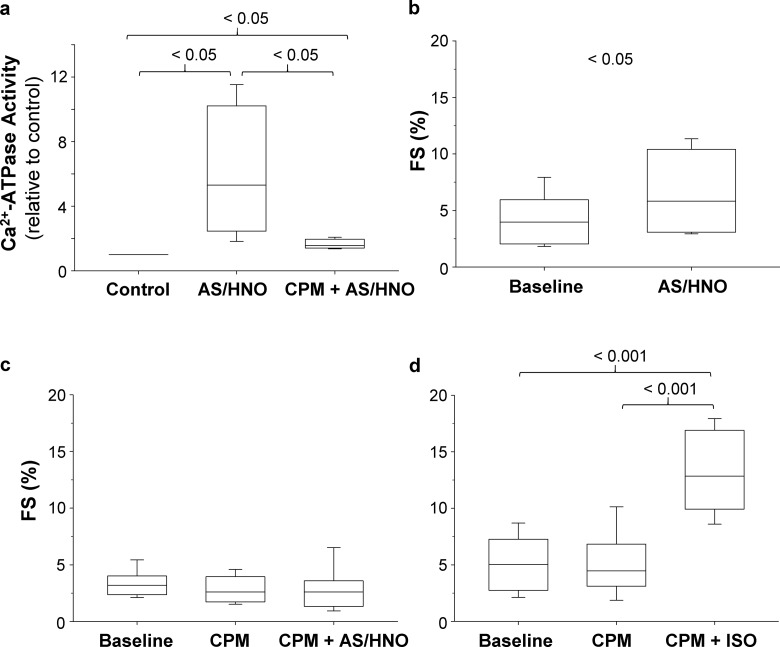
Nitroxyl (HNO) positively modulates myocardial function by accelerating Ca2+ reuptake into the sarcoplasmic reticulum (SR). HNO-induced enhancement of myocardial Ca2+ cycling and function is due to the modification of cysteines in the transmembrane domain of phospholamban (PLN), which results in activation of SR Ca2+-ATPase (SERCA2a) by functionally uncoupling PLN from SERCA2a. However, which cysteines are modified by HNO, and whether HNO induces reversible disulfides or single cysteine sulfinamides (RS(O)NH2) that are less easily reversed by reductants, remain to be determined. Using an 15N-edited NMR method for sulfinamide detection, we first demonstrate that Cys46 and Cys41 are the main targets of HNO reactivity with PLN. Supporting this conclusion, mutation of PLN cysteines 46 and 41 to alanine reduces the HNO-induced enhancement of SERCA2a activity. Treatment of WT-PLN with HNO leads to sulfinamide formation when the HNO donor is in excess, whereas disulfide formation is expected to dominate when the HNO/thiol stoichiometry approaches a 1:1 ratio that is more similar to that anticipated in vivo under normal, physiological conditions. Thus, 15N-edited NMR spectroscopy detects redox changes on thiols that are unique to HNO, greatly advancing the ability to detect HNO footprints in biological systems, while further differentiating HNO-induced post-translational modifications from those imparted by other reactive nitrogen or oxygen species. The present study confirms the potential of HNO as a signaling molecule in the cardiovascular system.
© 2019 Keceli et al.
Figures
Comment in
-
Tracking nitroxyl-derived posttranslational modifications of phospholamban in cardiac myocytes.J Gen Physiol. 2019 Jun 3;151(6):718-721. doi: 10.1085/jgp.201912342. Epub 2019 Apr 22. J Gen Physiol. 2019. PMID: 31010809 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
